Мотонейроны периферические (альфа- и гамма-мотонейроны)
Периферические
мотонейроны подразделяются на
альфа-мотонейроны и гамма-мотонейроны
(рис.
21.2 ).
Меньшие
по размеру гамма-мотонейроны
иннервируют интрафузальные
мышечные волокна .
Активация гамма-мотонейронов увеличивает
растяжение мышечных веретен, тем самым
облегчаясухожильные и
другие рефлексы, замыкающиеся через
альфа-мотонейроны.
Каждую
мышцу иннервирует несколько сотен
альфа-мотонейронов. В свою очередь,
каждый альфа-мотонейрон иннервирует
множество мышечных волокон — около
двадцати в наружных
мышцах глаза и
сотни в мышцах
конечностей и туловища .
В
нервно-мышечных синапсах
выделяется ацетилхолин .
Аксоны периферических
мотонейронов идут в составе черепных
нервов и передних
корешков спинного мозга .
На уровне межпозвоночных
отверстий передние
корешки и задние
корешкисливаются,
образуя спинномозговые
нервы .
Несколько соседних спинномозговых
нервов образуют сплетение, а затем
разветвляются на периферические
нервы .
Последние тоже неоднократно разветвляются
и иннервируют несколько мышц. Наконец,
аксон каждого альфа-мотонейрона образует
многочисленные разветвления, иннервируя
многие мышечные волокна.
Каждый
альфа-мотонейрон получает прямые
возбуждающие глутаматергические входы
откорковых
мотонейронов и
от чувствительных
нейронов ,
иннервирующих мышечные
веретена .
Возбуждающие влияния поступают также
к альфа- и гамма-мотонейронам от двигательных
ядер ствола мозга и вставочных
нейронов спинного мозга —
как по прямым путям, так и с переключениями.
Прямое
постсинаптическое торможение
альфа-мотонейронов осуществляют клетки
Реншоу —
вставочные глицинергические
нейроны .
Непрямое пресинаптическое торможение
альфа-мотонейронов и непрямое
пресинаптическое торможение
гамма-мотонейронов обеспечивают
другие нейроны ,
образующие ГАМКергические синапсы на
нейронах задних
рогов .
Тормозное
действие на альфа- и гамма-мотонейроны
оказывают и другие вставочные нейроны
спинного мозга, а также двигательные
ядра ствола мозга .
Если
преобладают возбуждающие входы, группа
периферических мотонейронов активируется.
Вначале возбуждаются мелкие мотонейроны.
По мере того как сила сокращения мышцы
нарастает, частота их разрядов нарастает
и вовлекаются крупные мотонейроны. При
максимальном сокращении мышцы возбуждена
вся соответствующая группа мотонейронов.
Нейронные структуры и их свойства
Тела
чувствительных клеток вынесены за
пределы спинного мозга (рис.9.1.). Часть
из них располагается в спинальных
ганглиях. Это тела соматических
афферентов, иннервирующих в основном
скелетные мышцы. Другие находятся в
экстра- и интрамуральных ганглиях
автономной нервной системы и обеспечивают
чувствительность только внутренних
органов.
Чувствительные
клетки имеют один отросток, который
вскоре после отхождения от тела клетки
делится на две ветви.
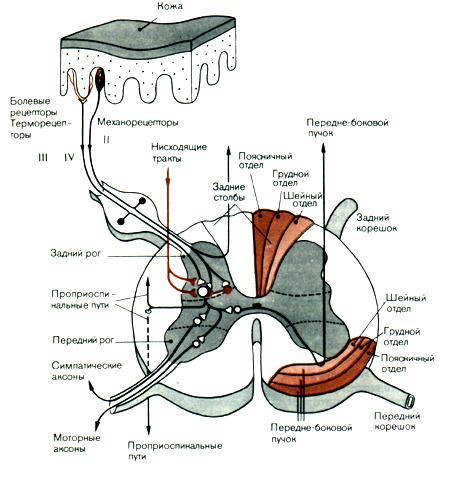
Рис.9.1.
Поперечное сечение спинного мозга и
связи кожных афферентов в спинном мозгу.
Одна
из них проводит возбуждение от рецепторов
к телу клетки, другая — от тела нервной
клетки к нейронам спинного или головного
мозга. Распространение возбуждения из
одной ветви в другую может происходить
без участия тела клетки.
Нервные
волокна чувствительных клеток по
скорости проведения возбуждения и
диаметру классифицируют на А-, В- и
С-группы. Толстые миелинизированные А-волокна с
диаметром от 3 до 22 мкм и скоростью
проведения возбуждения от 12 до 120 м/с
разбиваются еще на подгруппы: альфа—
волокна от мышечных рецепторов, бета —
от тактильных рецепторов и
барорецепторов,дельта —
от терморецепторов, механорецепторов,
болевых рецепторов. К волокнам
группы В относят
миелиновые отростки средней толщины
со скоростью проведения возбуждения
3-14 м/с. По ним в основном передается
ощущение боли. К афферентным волокнам
типа С относится
большинство безмиелиновых волокон
толщиной не более 2 мкм и скоростью
проведения до 2 м/с. Это волокна от
болевых, хемо- и некоторых механорецепторов.
Сам
спинной мозг в целом содержит, например,
у человека примерно 13 млн. нейронов. Из
их общего числа только около 3% составляют
эфферентные, двигательные или моторные
нейроны, а остальные 97 % приходятся на
долю вставочных, или интернейронов.
Двигательные нейроны являются выходными
клетками спинного мозга. Среди них
различают альфа- и гамма-мотонейроны,
а также преганглионарные нейроны
автономной нервной системы.
Альфа-мотонейроны осуществляют
передачу скелетным мышечным волокнам
сигналов, выработанных в спинном мозгу.
Аксоны каждого мотонейрона многократно
делятся, и, таким образом, каждый из них
охватывает своими терминалями до сотни
мышечных волокон, образуя в совокупности
с ними двигательную
единицу.
В свою очередь, несколько мотонейронов,
иннервирующих одну и ту же мышцу,
образуют мотонейронный
пул,
В его состав могут входить мотонейроны
нескольких соседних сегментов. В связи
с тем что возбудимость мотонейронов
пула неодинакова, при слабых раздражениях
возбуждается только часть из них. Это
влечет за собой сокращение лишь части
мышечных волокон. Другие моторные
единицы, для которых это раздражение
является подпороговым, тоже реагируют,
хотя их реакция выражается лишь в
деполяризации мембраны и повышении
возбудимости. С усилением раздражения
они еще больше вовлекаются в реакцию,
и, таким образом, все двигательные
единицы пула участвуют в рефлекторном
ответе.
Максимальная
частота воспроизведения ПД в
альфа-мотонейроне не превышает 200-300
имп/с. Вслед за ПД, амплитуда которого
составляет 80-100 мВ, возникает следовая
гиперполяризациядлительностью
от 50 до 150 мс. По частоте импульсации и
выраженности следовой гиперполяризации
мотонейроны разделяют на две группы:
фазические и тонические. Особенности
их возбуждения коррелируют с функциональными
свойствами иннервируемых мышц.
Фазически-ми мотонейронами иннервируются
более быстрые, «белые» мышцы,
тоническими — более медленные, «красные»
.
В
организации функции альфа-мотонейронов
важным звеном является наличие системы
отрицательной обратной связи,
образованной аксонными коллатералями
и специальными тормозными вставочными
нейронами — клетками Реншоу. Своими
возвратными тормозными влияниями они
могут охватывать большие группы
мотонейронов, обеспечивая, таким образом,
интеграцию процессов возбуждения и
торможения.
Гамма-мотонейроны иннервируют
интрафузальные (внутриверетенные)
мышечные волокна. Они разряжаются с
более низкой частотой, и следовая
гиперполяризация у них выражена слабее,
чем у альфа-мотонейронов. Их функциональное
значение сводится к сокращению
интрафузальных мышечных волокон, не
приводящему, однако, к появлению
двигательного ответа. Возбуждение этих
волокон сопровождается изменением
чувствительности их рецепторов к
сокращению или расслаблению экстрафузальных
мышечных волокон.
Нейроны
автономной нервной системы составляют
особую группу клеток. Тела симпатических
нейронов,
аксоны которых являются преганглионарными
волокнами, располагаются в
интермедиолатеральном ядре спинного
мозга. По своим свойствам они относятся
к группе B-волокон. Характерной особенностью
их функционирования является низкая
частота свойственной им постоянной
тонической импульсной активности. Одни
из этих волокон участвуют в поддержании
сосудистого тонуса, другие — обеспечивают
регуляцию висцеральных эффекторных
структур (гладкой мускулатуры
пищеварительной системы, железистых
клеток).
Тела парасимпатических
нейронов образуют
крестцовые парасимпатические ядра. Они
располагаются в сером веществе крестцовых
сегментов спинного мозга. Для многих
из них характерна фоновая импульсная
активность, частота которой возрастает
по мере повышения давления в мочевом
пузыре. При раздражении висцеральных
тазовых афферентных волокон в этих
эфферентных клетках регистрируется
вызванный разряд, характеризующийся
чрезвычайно большим латентным периодом.
К вставочным,
или интернейронам,
спинного мозга относятся нервные клетки,
аксоны которых не выходят за его пределы.
В зависимости от хода отростков различают
собственно спинальные и проекционные. Спинальные
интернейроны ветвятся
в пределах нескольких смежных сегментов,
образуя внутрисегментарные и
межсегментарные связи. Наряду с ними
существуют интернейроны, аксоны которых
проходят через несколько сегментов или
даже из одного отдела спинного мозга в
другой. Их аксоны образуют собственные
пучки спинного мозга.
К проекционным
интернейронам относятся
клетки, длинные аксоны которых формируют
восходящие пути спинного мозга. На
каждом интернейроне расположено в
среднем около 500 синапсов. Синаптические
влияния в них опосредуются через ВПСП
и ТПСП, суммация которых и достижение
критического уровня приводят к
возникновению распространяющегося ПД.
Соседние файлы в папке Нейрофизиология
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
| Альфа-мотонейрон | |
|---|---|
 Альфа-мотонейрон происходит из базальной пластинки (базальной пластинки) развивающийся эмбрион. Альфа-мотонейрон происходит из базальной пластинки (базальной пластинки) развивающийся эмбрион. |
|
| Идентификаторы | |
| NeuroLex ID | sao1154704263 |
| TH | H2.00.01.0.00008 |
| FMA | 83664 |
| Анатомические термины нейроанатомии [редактировать в Викиданных ] |
Альфа (α) моторные нейроны (также называемые альфа-мотонейронами ), большие, многополярные нижние моторные нейроны ствола мозга и спинной мозг. Они иннервируют экстрафузальные мышечные волокна скелетных мышц и непосредственно ответственны за начало их сокращения. Альфа-мотонейроны отличаются от гамма-мотонейронов, которые иннервируют интрафузальные мышечные волокна мышечных веретен.
, тогда как их клеточные тела находятся в центральная нервная система (ЦНС), α мотонейроны также считаются частью соматической нервной системы — ветви периферической нервной системы (ПНС) — потому что их аксоны проходят на периферию, чтобы иннервировать скелетные мышцы.
Альфа-мотонейрон и мышечные волокна, которые он иннервирует, являются моторной единицей. Пул моторных нейронов содержит тела всех альфа-мотонейронов, участвующих в сокращении одной мышцы.
Содержание
- 1 Местоположение
- 1.1 Ствол мозга
- 1.2 Спинной мозг
- 2 Развитие
- 3 Связь
- 3.1 Афферентный вход
- 3.2 Эфферентный выход
- 4 Сигнализация
- 5 Клиническая значимость
- 6 См. Также
- 7 Ссылки
- 8 Внешние ссылки
Местоположение
Альфа-мотонейроны (α-МН), иннервирующие голову и шея находятся в стволе мозга ; остальные α-MN иннервируют остальную часть тела и находятся в спинном мозге. В спинном мозге больше α-МН, чем в стволе мозга, поскольку количество α-МН прямо пропорционально степени контроля мелкой моторики в этой мышце. Например, мышцы одного пальца имеют больше α-MN на волокно и больше α-MN в целом, чем мышцы четырехглавой мышцы, что позволяет более точно контролировать силу, прилагаемую пальцем.
Как правило, α-МН на одной стороне ствола или спинного мозга иннервируют мышцы на той же стороне тела. Исключение составляет блокированное ядро в стволе мозга, которое иннервирует верхнюю косую мышцу глаза на противоположной стороне лица.
Ствол мозга
В стволе мозга α-MN и другие нейроны находятся в кластерах клеток, называемых ядрами, некоторые из которых содержат тела клеток. нейронов черепных нервов. Не все ядра черепных нервов содержат α-MN; те, что есть, являются двигательными ядрами, а другие — сенсорными ядрами. Моторные ядра находятся по всему стволу мозга — мозговому веществу, мосту и среднему мозгу — и по причинам развития находятся около средней линии ствола мозга.
Как правило, двигательные ядра, расположенные выше в стволе мозга (т.е. более ростральные), иннервируют мышцы, расположенные выше на лице. Например, глазодвигательное ядро содержит α-МН, которые иннервируют мышцы глаза, и находится в среднем мозге, самом ростральном компоненте ствола мозга. Напротив, подъязычное ядро , которое содержит α-МН, иннервирующие язык, находится в продолговатом мозге, наиболее каудальной (то есть в нижней части) структур ствола мозга.
Спинной мозг
 Кортикоспинальный тракт является одним из основных нисходящих путей от головного мозга к α-МН спинного мозга.
Кортикоспинальный тракт является одним из основных нисходящих путей от головного мозга к α-МН спинного мозга.
В спинном мозге α-МН расположены в сером веществе, которое образует вентральный рог. Эти α-МН обеспечивают двигательный компонент спинномозговых нервов, которые иннервируют мышцы тела.
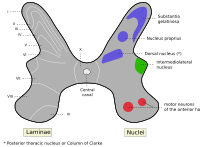 Альфа-мотонейроны расположены в ламине IX в соответствии с системой пластинок Рекседа..
Альфа-мотонейроны расположены в ламине IX в соответствии с системой пластинок Рекседа..
Как и в стволе мозга, более высокие сегменты спинного мозга содержат альфа-МН, которые иннервируют мышцы, расположенные выше на теле. Например, двуглавая мышца плеча, мышца руки, иннервируется α-МН в сегментах C5, C6 и C7 спинного мозга, которые находятся рострально в спинном мозге. С другой стороны, икроножная мышца, одна из мышц ноги, иннервируется α-МН в сегментах S1 и S2, которые находятся каудально в спинном мозге.
Альфа-мотонейроны расположены в определенной области серого вещества спинного мозга. Эта область обозначена lamina IX в системе пластинок Rexed, которая классифицирует области серого вещества на основе их цитоархитектуры. Lamina IX располагается преимущественно в медиальной части вентрального рога, хотя есть некоторый вклад в lamina IX за счет набора двигательных нейронов, расположенных более латерально. Как и другие области спинного мозга, клетки в этой пластинке соматотопически организованы, что означает, что положение нейронов в спинном мозге связано с тем, какие мышцы они иннервируют. В частности, α-МН в медиальной зоне ламины IX имеют тенденцию иннервировать проксимальные мышцы тела, в то время как те, что в боковой зоне, имеют тенденцию иннервировать более дистальные мышцы. Аналогичная соматотопия связана с α-МН, которые иннервируют мышцы-сгибатели и разгибатели: α-МН, которые иннервируют сгибатели, как правило, располагаются в дорсальной части пластинки IX; те, что иннервируют разгибатели, как правило, расположены более вентрально.
Развитие
 Под воздействием белка sonic hedgehog, показанного здесь, клетки пластинки дна развивающегося спинного мозга дифференцируются в альфа-мотонейроны.
Под воздействием белка sonic hedgehog, показанного здесь, клетки пластинки дна развивающегося спинного мозга дифференцируются в альфа-мотонейроны.
Альфа-мотонейроны берут начало в базальной пластинке, вентральной части нервной трубки развивающегося эмбриона. Sonic hedgehog (Shh) секретируется ближайшей хордой и другими вентральными структурами (например, пластиной дна ), создавая градиент высокой концентрации Shh в базальная пластинка и менее концентрированная Shh в крыловой пластине. Под влиянием Shh и других факторов некоторые нейроны базальной пластинки дифференцируются в α-MN.
Как и другие нейроны, α-МН посылают аксональные проекции для достижения своей цели экстрафузальных мышечных волокон через наведение аксонов, процесс частично регулируется нейротрофическими факторами, высвобождаемыми целевыми мышечными волокнами. Нейротрофические факторы также гарантируют, что каждое мышечное волокно иннервируется соответствующим количеством α-МН. Как и большинство типов нейронов в нервной системе, α-MN более многочисленны на раннем этапе развития, чем во взрослом возрасте. Мышечные волокна секретируют ограниченное количество нейротрофических факторов, способных поддерживать только часть α-МН, которые первоначально проецируются в мышечные волокна. Те α-МН, которые не получают достаточного количества нейротрофических факторов, будут подвергаться апоптозу, форме запрограммированной гибели клеток.
Поскольку они иннервируют многие мышцы, некоторые кластеры α-МН получают высокие концентрации нейротрофических факторы и выживают на этой стадии отсечения нейронов. Это верно для α-МН, иннервирующих верхние и нижние конечности: эти α-МН образуют большие столбцы клеток, которые способствуют шейному и поясничному увеличению спинного мозга. Помимо получения нейротрофических факторов от мышц, α-МН также секретируют ряд трофических факторов для поддержки мышечных волокон, которые они иннервируют. Пониженный уровень трофических факторов способствует атрофии мышц, которая следует за поражением α-МН.
Связность
Как и другие нейроны, нижние двигательные нейроны имеют как афферентные (входящие), так и эфферентные (исходящие) связи. Альфа-мотонейроны получают входные данные от ряда источников, включая верхние мотонейроны, сенсорные нейроны и интернейроны. Первичный выход α-МН — в экстрафузальные мышечные волокна. Эта афферентная и эфферентная связь необходима для достижения скоординированной мышечной активности.
Афферентный вход
| происхождение UMN | цель α-MN | Название тракта |
|---|---|---|
| Кора головного мозга | Ствол мозга | Кортикоядерный тракт |
| Кора головного мозга | Спинной мозг | Кортикоспинальный тракт |
| Красное ядро | Спинной мозг | Руброспинальный тракт |
| Вестибулярные ядра | Спинной мозг | Вестибулоспинальный тракт |
| Тектум среднего мозга | Спинной мозг | Тектоспинальный тракт |
| Ретикулярная формация | Спинной мозг | Ретикулоспинальный тракт |
Верхние двигательные нейроны (UMNs) посылают входные данные в α-MN несколькими путями, включая (но не ограничиваясь) кортикоядер, кортикоспинальный и руброспинальный тракт. Кортикоядерный и кортикоспинальный тракты обычно встречаются при исследованиях взаимодействия верхних и нижних мотонейронов при контроле произвольных движений.
кортиконуклеарный тракт назван так потому, что он соединяет кору головного мозга с ядрами черепных нервов. (Кортиконуклеарный тракт также называется кортикобульбарным трактом, поскольку мишень в стволе мозга — продолговатый мозг — архаично называется «луковицей».) Именно по этому пути верхние мотонейроны спускаются из коры. и синапс на α-МН ствола мозга. Точно так же UMN коры головного мозга напрямую контролируют α-MN спинного мозга через латеральный и вентральный кортикоспинальный тракты.
Сенсорный вход в α- MNs обширны и происходят из органов сухожилия Гольджи, мышечных веретен, механорецепторов, терморецепторов и других сенсорных нейронов. на периферии. Эти связи обеспечивают структуру нейронных цепей, лежащих в основе рефлексов. Существует несколько типов рефлекторных цепей, самый простой из которых состоит из одного синапса между сенсорным нейроном и α-МН. рефлекс коленного рефлекса является примером такого моносинаптического рефлекса.
Самый обширный вход в α-МН поступает от локальных интернейронов, которые являются наиболее многочисленным типом нейронов в спинном мозге. Среди их многих функций — синапсы интернейронов на α-MN для создания более сложных рефлекторных схем. Одним из типов интернейронов является клетка Реншоу.
Эфферентный выход
Альфа-мотонейроны посылают волокна, которые в основном синапсы на экстрафузальные мышечные волокна. Другие волокна синапса α-MNs на клетках Реншоу, то есть тормозящие интернейроны, которые синапсируют на α-MN и ограничивают его активность, чтобы предотвратить повреждение мышц.
Передача сигналов
Как и другие нейроны, α-MN передают сигналы в виде потенциалов действия, быстрых изменений электрической активности, которые распространяются от тела клетки к конец аксона . Чтобы увеличить скорость перемещения потенциалов действия, аксоны α-MN имеют большой диаметр и сильно миелинизированы как олигодендроцитами, так и шванновскими клетками. Олигодендроциты миелинизируют часть аксона α-MN, которая находится в центральной нервной системе (ЦНС), в то время как клетки Шванна миелинизируют часть, которая находится в периферической нервной системе (ПНС). Переход между ЦНС и ПНС происходит на уровне мягкой мозговой оболочки, самого внутреннего и наиболее тонкого слоя менингеальной ткани, окружающего компоненты ЦНС.
Аксон α-МН соединяется с его экстрафузальным мышечным волокном через нервно-мышечное соединение, специальный тип химического синапса, который отличается как по структуре, так и по функциям от химические синапсы, соединяющие нейроны друг с другом. Оба типа синапсов полагаются на нейротрансмиттеры для преобразования электрического сигнала в химический сигнал и обратно. Одно из их различий заключается в том, что синапсы между нейронами обычно используют глутамат или ГАМК в качестве нейромедиаторов, в то время как нервно-мышечные соединения используют исключительно ацетилхолин. Ацетилхолин ощущается никотиновыми рецепторами ацетилхолина на экстрафузальных мышечных волокнах, вызывая их сокращение.
Как и другие двигательные нейроны, α-MN названы в честь свойств их аксонов. Альфа-моторные нейроны имеют аксоны Aα, которые представляют собой большие калибра, сильно миелинизированные волокна, которые быстро проводят потенциалы действия. Напротив, гамма-мотонейроны имеют аксоны Aγ, которые представляют собой тонкие, слегка миелинизированные волокна, которые проводят медленнее.
Клиническое значение
 Полиомиелит, вызванный полиовирусом, обнаруженным здесь, связан с избирательной потерей клеток в вентральном роге спинного мозга, где расположены α-МН.
Полиомиелит, вызванный полиовирусом, обнаруженным здесь, связан с избирательной потерей клеток в вентральном роге спинного мозга, где расположены α-МН.
Повреждение α-МН является наиболее распространенным типом поражения нижних мотонейронов . Повреждение может быть вызвано, среди прочего, травмой, ишемией и инфекцией. Кроме того, некоторые заболевания связаны с избирательной потерей α-MN. Например, полиомиелит вызывается вирусом, который специфически поражает двигательные нейроны в вентральном роге спинного мозга и убивает их. Боковой амиотропный склероз аналогичным образом связан с избирательной потерей двигательных нейронов.
Паралич — один из наиболее выраженных эффектов повреждения α-МН. Поскольку α-MN обеспечивают единственную иннервацию экстрафузальных мышечных волокон, потеря α-MN эффективно разрывает связь между стволом и спинным мозгом и мышцами, которые они иннервируют. Без этой связи невозможно произвольное и непроизвольное (рефлекторное) управление мышцами. Произвольный мышечный контроль теряется, потому что α-МН передают произвольные сигналы от верхних мотонейронов к мышечным волокнам. Потеря непроизвольного контроля возникает в результате прерывания рефлекторных цепей, таких как тонический рефлекс растяжения. Следствием прерывания рефлекса является снижение мышечного тонуса, в результате чего. Другим следствием является угнетение глубоких сухожильных рефлексов, вызывающих гипорефлексию.
мышечную слабость и атрофию, которые также являются неизбежными последствиями поражений α-МН. Поскольку размер и сила мышц зависят от степени их использования, денервированные мышцы склонны к атрофии. Вторичной причиной мышечной атрофии является то, что денервированные мышцы больше не снабжаются трофическими факторами от α-МН, которые их иннервируют. Поражения альфа-мотонейрона также приводят к аномальным ЭМГ потенциалам (например,) и фасцикуляциям, причем последние представляют собой спонтанные непроизвольные сокращения мышц.
Заболевания, которые нарушают передачу сигналов между α-МН и экстрафузальными мышечными волокнами, а именно заболевания нервно-мышечного соединения, имеют признаки, аналогичные тем, которые возникают при заболевании α-МН. Например, миастения представляет собой аутоиммунное заболевание, которое препятствует передаче сигналов через нервно-мышечное соединение, что приводит к функциональной денервации мышц.
См. Также
- Бета-мотонейрон
- Экстрафузальное мышечное волокно
- Гамма-мотонейрон
- Внутривенное мышечное волокно
- Мышечное веретено
- клетка Реншоу
Литература
Внешние ссылки
- Поиск NIF — Alpha Motor Neuron через Neuroscience Information Framework
| Альфа мотонейрон | |
|---|---|
|
Альфа-мотонейроны происходят из базальная пластинка (базальная пластинка) развивающейся эмбрион. |
|
| Идентификаторы | |
| НейроЛекс Я БЫ | sao1154704263 |
| TH | H2.00.01.0.00008 |
| FMA | 83664 |
| Анатомические термины нейроанатомии
[редактировать в Викиданных ] |
Альфа (α) двигательные нейроны (также называемый альфа-мотонейроны), большие, многополярный нижние двигательные нейроны из мозговой ствол и спинной мозг. Они иннервируют экстрафузальные мышечные волокна из скелетные мышцы и несут прямую ответственность за инициирование их сокращение. Альфа-мотонейроны отличаются от гамма мотонейроны, которые иннервируют интрафузальные мышечные волокна из мышечные веретена.
Пока их клеточные тела находятся в Центральная нервная система (ЦНС), α мотонейроны также считаются частью соматическая нервная система — филиал периферическая нервная система (PNS) — потому что их аксоны распространяться на периферию, чтобы иннервировать скелетные мышцы.
Альфа-мотонейрон и мышечные волокна, которые он иннервирует, представляют собой моторный блок. А пул моторных нейронов содержит тела всех альфа-мотонейронов, участвующих в сокращении одной мышцы.
Место расположения
Альфа-мотонейроны (α-МН), иннервирующие голова и шея находятся в мозговой ствол; остальные α-МН иннервируют остальную часть тела и находятся в спинной мозг. В спинном мозге больше α-МН, чем в стволе мозга, поскольку количество α-МН прямо пропорционально степени контроля мелкой моторики в этой мышце. Например, в мышцах одного пальца больше α-МН на одно волокно и в целом больше α-МН, чем в мышцах квадрицепс, что позволяет более точно контролировать силу, прилагаемую пальцем.
Как правило, альфа-МН на одной стороне ствола или спинного мозга иннервируют мышцы на той же стороне тела. Исключением является трохлеарное ядро в стволе мозга, который иннервирует верхняя косая мышца глаза на противоположной стороне лица.
Мозговой ствол
В стволе мозга α-МН и другие нейроны находятся в кластерах ячеек, называемых ядра, некоторые из которых содержат тела нейронов, принадлежащих к черепные нервы. Не все ядра черепных нервов содержат α-МН; те, которые делают это двигательные ядра, а другие сенсорные ядра. Моторные ядра находятся по всему стволу мозга.мозговое вещество, мосты, и средний мозг — и по причинам развития обнаруживаются около средней линии ствола мозга.
Как правило, двигательные ядра, расположенные выше в стволе мозга (т.е. более ростральные), иннервируют мышцы, расположенные выше на лице. Например, глазодвигательное ядро содержит α-МН, которые иннервируют мышцы глаза, и находится в среднем мозге, самом ростральном компоненте ствола мозга. Напротив, подъязычное ядро, который содержит α-МН, иннервирующие язык, находится в мозговом веществе, самом каудальном (то есть по направлению к основанию) структур ствола мозга.
Спинной мозг
В спинном мозге α-МН расположены внутри серое вещество что формирует брюшной рог. Эти α-MN обеспечивают двигательный компонент позвоночные нервы которые иннервируют мышцы тела.
Как и в стволе головного мозга, более высокие сегменты спинного мозга содержат α-МН, которые иннервируют мышцы выше на теле. Например, двуглавая мышца плеча, мышца руки, иннервируется α-МН в сегментах C5, C6 и C7 спинного мозга, которые находятся рострально в спинном мозге. С другой стороны, икроножная мышца, одна из мышц ноги, иннервируется α-МН в сегментах S1 и S2, которые расположены каудально в спинном мозге.
Альфа-мотонейроны расположены в определенной области серого вещества спинного мозга. Этот регион обозначен пластиной IX в Система пластин Rexed, который классифицирует области серого вещества на основе их цитоархитектура. Lamina IX располагается преимущественно в медиальной части вентрального рога, хотя есть некоторый вклад в lamina IX за счет набора двигательных нейронов, расположенных более латерально. Как и в других областях спинного мозга, клетки этой пластинки соматотопически организованный, что означает, что положение нейронов в спинном мозге связано с тем, какие мышцы они иннервируют. В частности, α-МН в медиальной зоне ламины IX имеют тенденцию иннервировать проксимальные мышцы тела, в то время как те, что в боковой зоне, имеют тенденцию иннервировать более дистальные мышцы. Аналогичная соматотопия связана с α-МН, которые иннервируют мышцы-сгибатели и разгибатели: α-МН, которые иннервируют сгибатели обычно располагаются в дорсальной части IX пластинки; те, которые иннервируют разгибатели имеют тенденцию располагаться более вентрально.
Разработка
Под влиянием протеина звуковой еж, показанные здесь, ячейки плита пола развивающегося спинного мозга дифференцируются в альфа-мотонейроны.
Альфа-мотонейроны происходят из базальная пластинка, вентральная часть нервная трубка в развивающихся эмбрион. Соник ежик (Шшш) секретируется ближайшими нотохорда и другие вентральные структуры (например, плита пола ), устанавливая градиент высококонцентрированного Shh в базальной пластине и менее концентрированного Shh в сигнальная пластина. Под влиянием Шш и других факторов некоторые нейроны базальной пластинки различать в α-МНС.
Как и другие нейроны, α-MN посылают аксональный прогнозы для достижения своей цели экстрафузальные мышечные волокна через управление аксоном, процесс, частично регулируемый нейротрофические факторы высвобождается целевыми мышечными волокнами. Нейротрофические факторы также гарантируют, что каждое мышечное волокно иннервируется соответствующим количеством α-МН. Как и в случае с большинством типов нейронов в нервная система, α-МН более многочисленны в раннем развитии, чем во взрослом возрасте. Мышечные волокна секретируют ограниченное количество нейротрофических факторов, способных поддерживать только часть α-МН, которые первоначально проецируются в мышечные волокна. Те α-МН, которые не получают достаточного количества нейротрофических факторов, будут подвергаться апоптоз, форма запрограммированная гибель клеток.
Поскольку они иннервируют многие мышцы, некоторые кластеры α-МН получают высокие концентрации нейротрофических факторов и выживают на этой стадии отсечения нейронов. Это верно для α-MN, иннервирующих верхние и нижние конечности: эти α-MN образуют большие столбцы клеток, которые вносят вклад в шейный и поясничные увеличения спинного мозга. Помимо получения нейротрофических факторов от мышц, α-МН также секретируют ряд трофические факторы для поддержки мышечных волокон, которые они иннервируют. Пониженный уровень трофических факторов способствует атрофии мышц, которая следует за поражением α-МН.
Связь
Как и другие нейроны, нижние двигательные нейроны имеют как афферентные (входящие), так и эфферентные (исходящие) связи. Альфа-мотонейроны получают входные данные от ряда источников, включая верхние двигательные нейроны, сенсорные нейроны, и интернейроны. Первичный выход α-MN должен экстрафузальные мышечные волокна. Эта афферентная и эфферентная связь необходима для достижения скоординированной мышечной активности.
Афферентный ввод
| UMN происхождение | α-MN мишень | Название тракта |
|---|---|---|
| Кора головного мозга | Мозговой ствол | Кортикоядерный тракт |
| Кора головного мозга | Спинной мозг | Кортикоспинальный тракт |
| Красное ядро | Спинной мозг | Руброспинальный тракт |
| Вестибулярные ядра | Спинной мозг | Вестибулоспинальный тракт |
| Тектум среднего мозга | Спинной мозг | Тектоспинальный тракт |
| Ретикулярная формация | Спинной мозг | Ретикулоспинальный тракт |
Верхние двигательные нейроны (UMN) отправляют входные данные в α-MN несколькими путями, включая (но не ограничиваясь) кортикоядерный, кортикоспинальный, и руброспинальные тракты. Кортикоядерный и кортикоспинальный тракты обычно встречаются при исследованиях взаимодействия верхних и нижних мотонейронов при контроле произвольных движений.
В кортикоядерный тракт назван так потому, что соединяет кора головного мозга к ядра черепных нервов. (Кортикоядерный тракт также называют кортикобульбарный тракт, как цель в стволе мозга, мозговое вещество — архаично называется «луковица».) Именно по этому пути верхние мотонейроны спускаются из коры и синапс на α-МН ствола мозга. Точно так же UMNs коры головного мозга находятся под прямым контролем α-MNs спинной мозг через боковой и вентральные кортикоспинальные тракты.
Сенсорный вклад в α-МН обширен и берет свое начало в Органы сухожилия Гольджи, мышечные веретена, механорецепторы, терморецепторы, и другие сенсорные нейроны на периферии. Эти связи обеспечивают структуру нейронных цепей, лежащих в основе рефлексы. Существует несколько типов рефлекторных цепей, самый простой из которых состоит из одного синапса между сенсорным нейроном и α-МН. В коленный рефлекс является примером такого моносинаптического рефлекса.
Наиболее обширный вклад в α-MN поступает от местных интернейроны, которые являются наиболее многочисленным типом нейронов в спинной мозг. Среди их многих функций — синапсы интернейронов на α-MN для создания более сложных рефлекторных схем. Одним из типов интернейронов является Ячейка Реншоу.
Эфферентный выход
Альфа-мотонейроны посылают волокна, которые в основном работают с синапсами. экстрафузальные мышечные волокна. Другие волокна синапса α-MNs на Клетки Реншоу, т.е. тормозящее интернейроны этот синапс на α-МН и ограничивает его активность, чтобы предотвратить повреждение мышц.
Сигнализация
Как и другие нейроны, α-MN передают сигналы как потенциалы действия, быстрые изменения электрической активности, происходящие от Тело клетки до конца аксон. Чтобы увеличить скорость перемещения потенциалов действия, аксоны α-MN имеют большой диаметр и миелинизированный обоими олигодендроциты и Шванновские клетки. Олигодендроциты миелинизируют часть аксона α-MN, лежащую в Центральная нервная система (ЦНС), тогда как клетки Шванна миелинизируют часть, лежащую в периферическая нервная система (ПНС). Переход между ЦНС и ПНС происходит на уровне pia mater, самый внутренний и самый тонкий слой менингеальная ткань окружающие компоненты ЦНС.
Аксон α-МН соединяется с экстрафузальным мышечным волокном через нервномышечное соединение, специализированный вид химический синапс который отличается как по структуре, так и по функциям от химических синапсов, соединяющих нейроны друг с другом. Оба типа синапсов полагаются на нейротрансмиттеры к преобразовывать электрический сигнал в химический сигнал и обратно. Одно из их различий заключается в том, что синапсы между нейронами обычно используют глутамат или же ГАМК как их нейротрансмиттеры, в то время как нервно-мышечный узел использует ацетилхолин исключительно. Ацетилхолин воспринимается никотиновые рецепторы ацетилхолина на экстрафузальные мышечные волокна, вызывая их сокращение.
Как и другие двигательные нейроны, α-MN названы в честь свойств их аксоны. Альфа-мотонейроны имеют Аксоны Aα, которые являются большимикалибр, сильно миелинизированный волокна, которые проводят потенциалы действия быстро. Напротив, гамма мотонейроны имеют Аксоны Aγ, которые представляют собой тонкие, слегка миелинизированные волокна, проводящие медленнее.
Клиническое значение
Повреждение α-МН — наиболее распространенный тип нижних мотонейронов. поражение. Ущерб может быть вызван травма, ишемия, и инфекционное заболевание, среди прочего. Кроме того, некоторые заболевания связаны с избирательной потерей α-MN. Например, полиомиелит вызвано вирус который нацеливается и убивает мотонейроны в вентральном роге спинного мозга. Боковой амиотропный склероз аналогично связано с избирательной потерей двигательных нейронов.
Паралич является одним из наиболее выраженных эффектов повреждения α-МН. Поскольку α-МН обеспечивают единственную иннервацию экстрафузальные мышечные волокна потеря α-МН эффективно разрывает связь между стволом и спинным мозгом и мышцами, которые они иннервируют. Без этой связи невозможно произвольное и непроизвольное (рефлекторное) управление мышцами. Произвольный мышечный контроль теряется, потому что α-МН передают произвольные сигналы от верхних двигательных нейронов мышечным волокнам. Потеря непроизвольного контроля возникает в результате прерывания рефлекторные цепи например тоник рефлекс растяжения. Следствием прерывания рефлекса является то, что мышечный тонус уменьшается, в результате вялый парез. Еще одно последствие — депрессия глубокие сухожильные рефлексы, вызывая гипорефлексия.
Слабость мышц и атрофия также являются неизбежными последствиями поражений α-МН. Поскольку размер и сила мышц зависят от степени их использования, денервированные мышцы склонны к атрофии. Вторичной причиной мышечной атрофии является то, что денервированные мышцы больше не снабжаются трофическими факторами от α-МН, которые их иннервируют. Поражения альфа-мотонейронов также приводят к аномальным ЭМГ потенциалы (например, потенциалы фибрилляции ) и фасцикуляции причем последнее является спонтанным непроизвольным сокращением мышц.
Заболевания, при которых нарушается передача сигналов между α-МН и экстрафузальными мышечными волокнами, а именно: заболевания нервно-мышечного узла имеют признаки, аналогичные тем, которые возникают при болезни α-МЯ. Например, миастения является аутоиммунное заболевание что предотвращает передачу сигналов через нервномышечное соединение, что приводит к функциональной денервации мышц.
Смотрите также
- Бета мотонейрон
- Экстрафузальное мышечное волокно
- Гамма двигательный нейрон
- Интрафузальное мышечное волокно
- Мышечное веретено
- Ячейка Реншоу
Рекомендации
- Джон А. Кирнан (2005). Человеческая нервная система Барра: анатомическая точка зрения (8-е изд.). Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 0-7817-5154-3.
- Дуэйн Э. Хейнс (2004). Нейроанатомия: Атлас структур, сечений и систем (6-е изд.). Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 0-7817-4677-9.
внешняя ссылка
- Поиск NIF — Alpha Motor Neuron через Информационная структура по неврологии
Functional Neural Transplantation III
Peter H. Chipman, … Victor F. Rafuse, in Progress in Brain Research, 2012
Abstract
Alpha motor neurons (also known as lower or skeletal motor neurons) have been studied extensively for over 100 years. Motor neurons control the contraction of skeletal muscles and thus are the final common pathway in the nervous system responsible for motor behavior. Muscles become paralyzed when their innervating motor neurons die because of injury or disease. Motor neuron diseases (MNDs), such as Amyotrophic Lateral Sclerosis, progressively destroy motor neurons until those inflicted succumb to the illness due to respiratory failure. One strategy being explored to study and treat muscle paralysis due to motor neuron loss involves deriving surrogate motor neurons from pluripotent stem cells. Guided by decades of research on the development of the spinal cord, recent advances in neurobiology have shown that functional motor neurons can be derived from mouse and human embryonic stem (ES) cells. Furthermore, ES cell-derived motor neurons restore motor behavior when transplanted into animal models of motor dysfunction. The recent discovery that mouse and human motor neurons can be derived from induced pluripotent stem (iPS) cells (i.e., somatic cells converted to pluripotency) has set the stage for the development of patient-specific therapies designed to treat movement disorders. Indeed, there is now hope within the scientific community that motor neurons derived from pluripotent stem cells will be used to treat MNDs through cell transplantation and/or to screen molecules that will prevent motor neuron death. In this chapter, we review the journey that led to the generation of motor neurons from ES and iPS cells, how stem cell-derived motor neurons have been used to treat/study motor dysfunction, and where the technology will likely lead to in the future.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780444595447000159
The nervous system
Nikolai Bogduk, in Anatomy and Human Movement (Second Edition), 1994
Motor nerve endings
Alpha-motor neurons form endings designed to deliver stimuli to extrafusal muscle fibres, and similar but smaller endings are formed by γ-motor neurons on intrafusal fibres. The junction between a motor neuron and a muscle cell involves elaboration of both the neuron and the muscle cell and of certain surrounding tissues, the entire complex being known as a motor endplate.
Structurally and functionally, neuromuscular junctions resemble synapses (see Fig. 7.3). As a motor axon approaches its target muscle cell it loses its myelin sheath and forms a flattened expansion that is applied to the surface of the muscle membrane. Once applied, the expansion is covered by a Schwann cell sheath which insulates the neuromuscular junction from the external environment. That portion of the muscle cell to which the nerve is apposed is modified by forming a flattened bump on its surface. This is formed by the focal accumulation of cytoplasm (sarcoplasm) and organelles within the muscle cell that raises its membrane underneath the nerve terminal. That part of the muscle cell membrane applied to the nerve cell is known as the postjunctional membrane (Fig. 7.49). Reciprocally, that part of the nerve cell membrane applied to the muscle cell is known as the prejunctional membrane.

Fig. 7.49. The microscopic structure of motor end-plates.
The postjunctional membrane forms troughs which lodge reciprocal folds of the prejunctional membrane. Within the troughs, the membranes of the nerve cell and muscle cell are separated by a synaptic cleft measuring about 20-50 nm which is filled with an amorphous ground substance. In the floor of the trough, the postjunctional membrane is thrown into further folds known as secondary synaptic clefts which increase the surface area of the receptive muscle membrane (Fig. 7.49).
The terminal expansion of the motor neuron contains vesicles filled with the transmitter substance acetylcholine. When an action potential arrives at the nerve terminal, the vesicles release the acetylcholine through the presynaptic membrane into the synaptic deft. The transmitter substance then flows across the cleft to react with molecular receptors on the postjunctional membrane which generates an action potential in the muscle membrane. This potential is then propagated into the muscle cell to cause contraction of the muscle fibres.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780750609708500137
Neuromotor Control of Speech
Wanda G. Webb PhD, CCC-SLP, in Neurology for the Speech-Language Pathologist (Sixth Edition), 2017
Alpha Motor Neurons
The alpha motor neuron (AMN) innervates the main fibers that cause muscle contraction. These fibers lie within the muscle and are called extrafusal fibers. Figure 6-6 illustrates the structure of a muscle fiber including the input from the AMN. The axon of each AMN branches to supply the fibers. An axon may supply only a few fibers, as in the case of a small muscle with precisely controlled contraction, or it may control several hundred fibers, as in the case of large muscles with strong, crude movements. This fact is consistent with what you saw on the homunculus illustrated in Figure 2-7. The oral musculature involved in speech and swallowing requires a much larger area on the motor strip because the innervation and control of these muscles requires the involvement of a vastly larger number of neurons to allow fine and precise motor control for these acts.
The alpha motor neuron supplies the trophic, or nutritional, factors, which direct differentiation of muscle fibers and keep the muscle healthy. These substances are called myotrophic factors. The motor neuron also supplies the acetylcholine that stimulates contraction of muscle.
Three types of extrafusal fibers can be differentiated in skeletal muscle. All muscles contain all three types of fibers, but all muscle fibers in one motor unit are of the same type. The type is determined by the myotrophic influences of the innervating neuron. The first of the three types of fibers are slow twitch (type I) fibers that contract slowly and are resistant to fatigue. These fibers predominate in sustaining postural activities, including standing. Fast twitch fibers (type IIx in humans) contract faster but fatigue more rapidly. They exert a more powerful force and are primarily found in superficial muscles. An intermediate fiber (Type IIA) has properties between the other two types in terms of speed of contraction and amount of force; it is considered a fast twitch fiber, however.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780323100274000063
Spinal Motor Neurons: Properties
H. Hultborn, B. Fedirchuk, in Encyclopedia of Neuroscience, 2009
Motor Neuronal Plasticity Following Training or Disuse
Do α-motor neurons, like muscle fibers, adapt to long-lasting increases or decreases in activity? There is much evidence to support the idea that motor activity and motor learning indeed involve neuronal plasticity at both the spinal and supraspinal levels of the central nervous system. However, it is only during the last several years that changes in the properties of α-motor neuron themselves in response to altered levels of physical activity have been directly assessed. The present evidence originates from experiments in which rats were subjected to forced endurance training or spontaneous running in exercise wheels to increase the motor activity or were subjected to body-weight support (i.e., suspension of the hindlimbs) for a couple of weeks to reduce the motor activity. Only a few examples of the described changes are mentioned here. With increased motor activity, the slow motor neurons showed more negative resting membrane potential as well as firing threshold. It was proposed that these changes may prevent depolarization, accommodation, and firing adaptation during prolonged activity. During inactivity, the resting membrane potential and firing threshold became more depolarized. The motor neurons become less excitable and more prone to accommodation and frequency adaptation. The signals mediating the adaptation of these (and other) motor neuron properties are not known at this time. However, it has been proposed that increased levels of neurotrophins originating from the exercised muscle, or other neurotrophic factors released by afferent activity, may induce appropriate changes in motor neuronal gene expression.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780080450469013425
Motor Control, Peripheral
N. Goyal, DA. Chad, in Encyclopedia of the Neurological Sciences (Second Edition), 2014
Sensory Neurons and the Simple Reflex Arc
The alpha motor neurons are influenced not only by upper motor neurons in the motor cortex and motor control neurons in the brainstem but also by sensory inputs from the periphery. In fact, the peripheral pathway that serves as the foundation for the generation of spontaneous muscle contraction in the resting state (also known as muscle tone) is the simple reflex arc. It is composed of a sensory or afferent limb (the heavily myelinated axon originating in the muscle spindle (known as Ia afferent fiber, the largest among all nerve fibers) whose unipolar cell body resides in the sensory or dorsal root ganglia); and a motor or efferent limb (the myelinated axon extending from the lower motor neuron). The afferent limb directly stimulates the efferent lower motor neuron via a single synapse; hence, the reflex arc is designated monosynaptic (Figure 9). The specialized unipolar structure of the sensory neuron cell body allows for its axon to bifurcate after leaving the cell body: one process passes to the muscle spindle in the periphery, whereas the other travels into the spinal cord to make its monosynaptic contact with the lower motor neuron.
With regard to the sensory innervation of the muscle spindle, the large-diameter myelinated axon (Ia) pierces the capsule of the spindle, loses its myelin sheath, and becomes a naked axon that winds spirally around the equatorial regions of the nuclear bag or chain portions of the intrafusal fibers (Figure 10). A slightly smaller diameter myelinated axon, arising from a smaller dorsal root ganglion neuron, also pierces the muscle spindle capsule, loses its myelin sheath, and forms a naked axon that branches terminally and ends as varicosities, resembling a spray of flowers. These endings are situated mainly on the nuclear chain fibers and some distance away from the equatorial region.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123851574006187
The Somatic Nervous System
Javier Cuevas, in xPharm: The Comprehensive Pharmacology Reference, 2007
The neuromuscular junction
Each alpha motoneuron innervates a motor unit, which is a population of muscle fibers. Although the number of muscle fibers innervated by a single motoneuron varies depending on the size and function of the muscle, there are about 150 muscle fibers per motor unit. While each muscle fiber receives input from a single motor neuron, motor units usually overlap such that 10–15 fibers from one motor unit are adjacent to similar bundles of a second motor unit. This permits motor units to work in unison and to contract in support of each other. Shown in Fig. 5A is a representation of four muscle fibers receiving innervation from a single motor neuron. Upon reaching the muscle fiber, the axon of the motoneuron divides into multiple terminal nerve twigs (Fig. 5B and 5D), all of which form synaptic contacts with the muscle fiber (Fig. 5C and 5E). These synaptic contacts are physically supported by both the basement membrane and overlapping Schwann cells (Fig. 5B and 5C). The presence of three cell types (neuron, glial and muscle) in the neuromuscular junction led to the term tripartite synapse.

Fig. 5. The motor unit and motor endplate. A: A single motoneuron innervates multiple muscle fibers forming a motor unit. B: Enlargement from (A) showing multiple synaptic contacts between a motoneuron and a single muscle fiber. C: Enlargement from (B) showing regions where synaptic activity occurs. Reprinted from Clinical Neuroanatomy and Related Neuroscience, from Fitzgerald, Copyright 2001, with permission from Elsevier. D: A methylene blue-stained muscle preparation showing several nerve twigs (T) branching from a single axon and innervating multiple muscle fibers. There is a bulbous swelling (the motor endplate, MEP) at the end of each twig at the site of connection with the muscle. E: Electron micrograph of a motor endplate showing the cell membrane of the muscle fiber arranged into a series of deep folds (junctional folds, JF) beneath which the sarcoplasm (S) contains numerous mitochondria (M). In the terminal swelling of the motor axon, neurosecretory granules (G), containing acetylcholine (ACh) and mitochondria are abundant. The terminal swelling of the axon is separated from the muscle cell membrane by a gap of 30–50 nm (the synaptic cleft, C), which includes the external lamina of the muscle. Reprinted from Human Histology,Stevens, Copyright 1996, with permission from Elsevier.
The synaptic contact between a motor neuron and skeletal muscle, or neuromuscular junction, is a highly specialized structure called the motor endplate (Fig. 5E). Neurosecretory vesicles that contain ACh are abundant on the presynaptic terminus of the motor axon. The synaptic cleft separating the neuron and nerve fiber at the motor endplate is 30–50 nm. The postsynaptic membrane of the muscle fiber contains numerous folds called junctional folds, and the apex of these junctional folds (membrane closes to the presynaptic terminus) exhibits pronounced optical density. These optically dense regions contain clusters of nicotinic acetylcholine receptors and associated proteins.
Depicted in Fig. 6 are some of the key molecules/components involved in structural organization and neurotransmission at the neuromuscular junction. The synaptic vesicles of the motor nerve terminal are released when an action potential reaches the presynaptic terminus. Activation of postsynaptic nicotinic receptors results in depolarization of the muscle fiber. If this depolarization is of sufficient magnitude, an action potential is evoked in the muscle fiber, and the resulting calcium influx and calcium release from intracellular stores activate the contraction machinery of the cell. Some of the proteins listed in Fig. 6 are also involved in antibody-mediated (autoimmune) or genetic disorders affecting motor control. For example, autoantibodies directed against nicotinic acetylcholine receptors on the skeletal muscle give rise to myasthenia gravis.

Fig. 6. Key components of neurotransmission at the neuromuscular junction. The synthesis of acetylcholine (ACh) is catalyzed by choline acetyl transferase (CHAT), after which the neurotransmitter is accumulated in synaptic vesicles, and released when an action potential reaches the nerve terminus and evokes calcium entry through voltage-gated calcium channels (VGCC). Upon release the ACh diffuses across the synaptic cleft and activates nicotinic ACh receptors (nAChRs) on the postsynaptic membrane. The nAChRs are clustered on the top of the junctional folds. Molecules that contribute to this clustering are and rapsyn. The clustering of nAChRs by these molecules is triggered by neural agrin (not shown). The depolarization initiated by ion flux through nAChRs activates voltage-gated sodium channels (VGNaCs) localized at the bottom of the folds and ultimately leads to elevation of intracellular calcium concentrations and muscle contraction. The action of ACh is terminated by the action of acetylcholinesterase, which is tethered in the synaptic basement membrane. Repolarization of the motor nerve terminal and the muscle fiber is mediated by opening of voltage-gated potassium channels (VGKCs) on the plasma membrane of both cells (only shown on motoneuron here). (Reproduced from (McConville and Vincent (2002)).
A variety of drugs exert their effects by influencing neurotransmission at the neuromuscular junction (Table 2).
Table 2. Drugs acting at the neuromuscular junction.
| Site of action | Agent Type | Drug Name |
|---|---|---|
| Presynaptic | Botulinum toxin | |
| Postsynaptic | Depolarizing Blocking Agents (nAChR agonist) | Succinylcholine |
| Non-Depolarizing Blocking Agents (nAChR antagonists) | Metocurine | |
| Pancuronium | ||
| Tubocurarine | ||
| Cholinesterase Inhibitors | Echothiophate | |
| Edrophonium | ||
| Malathion | ||
| Neostigmine | ||
| Physostigmine | ||
| Sarin | ||
| Soman |
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B978008055232360323X
The Somatic Nervous System☆
J. Cuevas, in Reference Module in Biomedical Sciences, 2015
The Neuromuscular Junction
Each α motor neuron innervates a population of muscle fibers termed a motor unit. The number of muscle fibers innervated by a single motoneuron varies depending on the size and function of the muscle, but averages about 150 muscle fibers per motor unit. While each muscle fiber receives input from a single motor neuron, motor units usually overlap whereby 10–15 fibers from one motor unit are adjacent to similar bundles of a second motor unit. This phenomenon permits motor units to work in unison and to contract in support of each other. Figure 5a shows a representation of four muscle fibers receiving innervation from a single motor neuron. Upon reaching the muscle fiber, the axon of the motoneuron divides into multiple terminal nerve twigs (Figure 5b and d), all of which form synaptic contacts with the muscle fiber (Figure 5c and e). These synaptic contacts are physically supported by both the basement membrane and overlapping Schwann cells (Figure 5b and c). The presence of three cell types (neuron, glial and muscle) in the neuromuscular junction led to the term tripartite synapse.

Figure 5. The motor unit and motor endplate. (a) A single motoneuron innervates multiple muscle fibers forming a motor unit. (b) Enlargement from (a) showing multiple synaptic contacts between a motoneuron and a single muscle fiber. (c) Enlargement from (b) showing regions where synaptic activity occurs. (Fitzgerald: Clinical Neuroanatomy and Related Neuroscience Figure 8.2). (d) A methylene blue-stained muscle preparation showing several nerve twigs (T) branching from a single axon and innervating multiple muscle fibers. There is a bulbous swelling (the motor endplate, MEP) at the end of each twig at the site of connection with the muscle. (e) Electron micrograph of a motor endplate showing the cell membrane of the muscle fiber thrown into a series of deep folds (junctional folds, JF) beneath which the sarcoplasm (S) contains numerous mitochondria (M). In the terminal swelling of the motor axon, neurosecretory granules (G), containing the transmitter substance acetylcholine (ACh), and mitochondria are abundant. The terminal swelling of the axon is separated from the muscle cell membrane by a gap of 30–50 nm (the synaptic cleft, C), which includes the external lamina of the muscle.
The synaptic contact between a motor neuron and skeletal muscle, or neuromuscular junction, is a highly specialized structure called the motor endplate. Figure 5e shows an electron micrograph of a motor endplate. On the presynaptic terminus of the motor axon neurosecretory vesicles that contain ACh are abundant. The synaptic cleft separating the neuron and nerve fiber at the motor endplate is 30–50 nm. The postsynaptic membrane of the muscle fiber contains numerous folds called junctional folds, and the apex of these junctional folds (membrane closes to the presynaptic terminus) exhibits pronounced optical density. These optically dense regions contain clusters of nicotinic ACh receptors and associated proteins.
Figure 6 depicts some of the key molecules/components involved in structural organization and neurotransmission at the neuromuscular junction. The synaptic vesicles of the motor nerve terminal are released when an action potential reaches the presynaptic terminus. Activation of postsynaptic nicotinic receptors results in depolarization of the muscle fiber. If this depolarization is of sufficient magnitude, an action potential will be evoked in the muscle fiber, and the resulting calcium influx and calcium release from intracellular stores will activate the contraction machinery of the cell. Some of the proteins listed in Figure 6 are also involved in antibody-mediated (autoimmune) or genetic disorders affecting motor control. For example, autoantibodies directed against nicotinic ACh receptors on the muscle give rise to myasthenia gravis.

Figure 6. Key component of neurotransmission at the neuromuscular junction: ACh is synthesized by choline acetyl transferase (CHAT), taken up into synaptic vesicles, and released when an action potential reaches the nerve terminus and evokes calcium entry through voltage-gated calcium channels (VGCC). ACh diffuses across the synaptic cleft and activates nicotinic ACh receptors (nAChRs) on the postsynaptic membrane. nAChRs are clustered on the top of the junctional folds. Two molecules that contribute to this clustering are muscle-specific kinase (MuSK) and rapsyn. The clustering of nAChRs by these molecules is triggered by neural agrin (not shown in this diagram). The depolarization initiated by ion flux through nAChRs activates voltage-gated sodium channels (VGNaCs) localized at the bottom of the folds and ultimately leads to elevation of intracellular calcium concentrations and muscle contraction. The action of ACh is terminated by its hydrolysis by AChesterase (AChE), which is tethered in the synaptic basement membrane. Repolarization of the motor nerve terminal and the muscle fiber is mediated by opening of voltage-gated potassium channels (VGKCs) that are found on the plasma membrane of both cells (only shown on motoneuron here).
Figure 1 from McConville J, Vincent A., Diseases of the neuromuscular junction. Curr Opin Pharmacol. 2002.
A variety of drugs exert their effects by influencing neurotransmission at the neuromuscular junction. Table 2 lists some of these agents and categorizes them as presynaptic or postsynaptic, based on their site of action. More information of these cholinergic synapses is provided in Neurotransmitters and Their Life Cycles and Neurotransmission.
Table 2. Drugs acting at the neuromuscular junction
| Site of action | Agent type | Drug name |
|---|---|---|
| Presynaptic | Botulinum toxin | |
| Postsynaptic | Depolarizing blocking agents (nAChR agonist) | Succinylcholine |
| Non-depolarizing blocking agents (nAChR antagonists) | Atracurium cisatracurium doxacurium Mivacurium pancuronium Pipecuronium Tubocurarine Vecuronium |
|
| Cholinesterase inhibitors | Echothiophate Edrophonium Malathion (malaoxona) Neostigmine Physostigmine Sarin Soman |
- a
- Active metabolite.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780128012383053642
Peripheral Nervous System
Andrei V. Krassioukov, in Encyclopedia of the Human Brain, 2002
V.C.2.a Neuromuscular Junctions in Skeletal Muscles
A single α motor neuron (located within the ventral horn of the spinal cord; see Fig. 1), with the muscle fibers that it innervates, is defined as the motor unit. As soon as the large myelinated axon of an α motor neuron enters a skeletal muscle, it branches many times. Each small branch terminates on muscle fibers at the motor end plate or neuromuscular junction (Fig. 11). The plasma membrane of the axon terminal (axolemma) is separated by the synaptic cleft (gap of 20–50 nm) from the plasma membrane of the muscle fiber (sarcolemma). The surface area of sarcolemma (postsynaptic membrane) at a motor end plate is thrown into numerous folds. These serve to increase the contact area of muscle to the naked axon (presynaptic membrane). The neurotransmitter, acetylcholine, is released from synaptic vesicles into the synaptic cleft when a nerve impulse reaches the motor end plate. Once the acetylcholine is released, it will stimulate receptor sites on the postsynaptic muscular membrane, causing the contraction of skeletal muscle fibers. The acetylcholine remains in contact with the postsynaptic membrane for a very short period of time (about 1 msec), and it is rapidly inactivated by the enzyme acetylcholinesterase (AChE). Acetylcholinesterase hydrolyzes acetylcholine into acetic acid and choline. The choline is taken up by the presynaptic terminals for the synthesis of new molecules of acetylcholine.

Figure 11. Motor end plate on skeletal muscle. Gold chloride method.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B0122272102002764
Strength and Reflexes
John P. Hammerstad, in Textbook of Clinical Neurology (Third Edition), 2007
Gamma Motor Neuron
Like the alpha motor neurons, the gamma motor neurons lie in the ventral horn of the spinal cord interspersed among the alpha motor neurons innervating the same muscle. Gamma motor neurons innervate the intrafusal muscle fibers of a specialized sensory organ, the muscle spindle.11 The muscle spindle consists of a small bag of muscle fibers that lie in parallel with the extrafusal skeletal muscle fibers. Therefore, when the muscle lengthens or shortens, the intrafusal muscle spindle fibers are stretched or relaxed correspondingly. The intrafusal fibers are surrounded by sensory nerve endings that become 1a afferents to the dorsal root ganglion. When the intrafusal fibers are stretched as the muscle is lengthened, the sensory fibers are activated, providing sensory feedback about the degree of lengthening that has occurred. When the muscle contracts and shortens, the intrafusal spindle fibers also shorten. If this were a completely passive system, the intrafusal fibers would relax and the sensory endings would become silent, providing no helpful feedback information about the state of the muscle. To prevent this inefficient circumstance, gamma motor neurons are activated, maintaining tension in the intrafusal fibers to continue to provide precise sensory information.
This phenomenon can be observed experimentally if alpha motor neurons are excited in isolation. When the muscle contracts, there is a pause in volleys from the 1a afferent. When the gamma motor neuron is excited at the same time, the afferent volleys do not pause. This simultaneous activation of alpha and gamma motor neurons during muscle contraction is called alpha‐gamma coactivation; it is an excellent example of sensory motor integration in the nervous system. By this means, innervation of the muscle spindle by an independent system of gamma motor neurons allows the central nervous system to adjust the sensitivity of the spindle and fine‐tune the information it receives (Fig. 15‐9).
Another sensory organ, the Golgi tendon organ, also conveys information about the state of the muscle. The Golgi tendon organs lie in the muscle tendon and, unlike the muscle spindle, are coupled in series with the extrafusal muscle fibers. Therefore, both passive stretch and active contraction of the muscle increase the tension of the tendon and activate the tendon organ receptor. In contrast to the activity of the muscle spindle, which depends on muscle length, the Golgi tendon organ conveys information about muscle tension. Together they convey precise information about the length, tension, velocity, and force of the muscle contraction, thus allowing greater precision of movement.12
Changes in muscle tone, defined as resistance to passive stretch of muscle, are an important feature of diseases of the motor system. At one time, an important component of normal resting muscle tone was thought to be the result of low‐level background alpha‐gamma coactivation. However, more recent studies in fully relaxed individuals indicate that the viscoelastic properties of muscle and tendons account for normal resting tone.13 Common experience indicates that resting muscle tone can vary considerably depending on the state of relaxation of the muscle as well as its viscoelastic properties. Some studies have shown that the passive viscoelastic properties of muscle may contribute to increased tone in patients with chronic spasticity and rigidity, but the most important determinants of pathological alterations in tone are the result of alterations in stretch reflexes.
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9781416036180100153
Spinal Cord
Gulgun Sengul, Charles Watson, in The Mouse Nervous System, 2012
Lamina 9
Lamina 9 contains large α-motor neurons whose axons innervate skeletal muscles, and small γ-motor neurons which innervate the muscle spindles. Lamina 9 also contains small interneurons and a small number of β-motor neurons. Lamina 9 neurons are characteristically cholinergic (Trifonov et al., 2009; Yato et al., 1997). Mouse lamina 9 neurons have also been shown to contain CGRP (Gibson et al., 1984b; Tie-Jun et al., 2001). In the rat, other substances that have been identified in motor neurons include calmodulin (Ren and Ruda, 1994), calcitonin gene-related peptide (Gibson et al., 1984), and dopamine (Holstege et al., 1996).
Read full chapter
URL:
https://www.sciencedirect.com/science/article/pii/B9780123694973100135
Краткое описание:
Библиографическая ссылка для цитирования: Сазонов В.Ф. Спинной мозг [Электронный ресурс] // Кинезиолог, 2009-2021: [сайт]. Дата обновления: 05.12.2021. URL: http://kineziolog.su/content/spinnoi-mozg (дата обращения: __.__.20__). _____________________________Отделы мозга, краткое описание спинного мозга, определение, схема его рефлекторных дуг.
Спинной мозг
Рисунок 1. Строение спинного мозга. Источник изображения: https://i08.fotocdn.net/s118/02fd10050bad7326/public_pin_l/2699904778.jpg
Рисунок 2. Строение спинного мозга: 1- Позвоночник; 2- Спинной мозг; 3- Суставной отросток; 4- Поперечный отросток; 5- Остистый отросток; 6- Место крепления ребра; 7- Тело позвонка; 8- Межпозвонковый диск; 9- Спинномозговой нерв; 10- Центральный канал спинного мозга; 11- Позвоночный нервный узел; 12- Мягкая оболочка; 13- Паутинная оболочка; 14- Твердая оболочка. Источник изображения: https://fit-baza.com/vegetativnaya-nervnaya-sistema-cheloveka/
Группы нервов:
I. Шейные нервы.
II. Грудные нервы.
III. Поясничные нервы.
IV. Крестцовые нервы.
V. Копчиковые нервы.
Головной мозг

ЦНС = отделы головного мозга + спинной мозг
1. Передний мозг.
2. Промежуточный мозг.


3. Средний мозг.
4. Варолиев мост — расширенная часть под средним мозгом.
5. Мозжечок (слева от него также красным цветом выделен Варолиев мост).

 6. Продолговатый мозг.
6. Продолговатый мозг.
7. Спинной мозг
Детали:
- шейное утолщение;
- пояснично-крестцовое утолщение;
- «конский хвост» — пучки нервов.
Спинной мозг
Введение
Угадайте, какая структура самая древняя в центральной нервной системе человека?
Конечно же, спинной мозг! Именно с него всё начиналось в эволюционном развитии мозга. Именно с него начинается развитие нервной системы у каждого человека, когда он находится ещё в состоянии зародыша. И именно со спинного мозга мы начинаем рассматривать строение и работу центральной нервной системы. Если мы хорошо поймём принципы устройства и работы спинного мозга, то нам легче будет понять и всё остальное.
Определение
Спинной мозг (по латыни Medulla spinalis) — это самая древняя хвостовая часть мозга, протянувшаяся в позвоночном канале вдоль тела и обеспечивающая управление мышцами и внутренними органами, а также связь с головным мозгом.
Спинной мозг — это самая филогенетически (эволюционно) древняя и онтогенетически (эмбрионально) ранняя структура центральной нервной системы, образовавшаяся из нервной трубки и имеющая метамерное строение. Функционально спинной мозг обеспечивает управление исполнительными органами (мышцами и железами), а также связь всей центральной нервной системы, включая головной мозг, с воспринимающими и исполнительными структурами организма. © Сазонов В.Ф., 2012. © kineziolog.bodhy.ru, 2012.
Спинной мозг — это «исполнительный отдел» ЦНС, почти все действия идут через него.
«Запоминалка» (метафора для памяти)
Спинной мозг — это «рабочая лошадка» ЦНС, и все остальные отделы мозга хотят на нём «ездить». Считаете сравнение слишком вольным? А как же тогда быть с тем, что у спинного мозга совершенно официально есть конский хвост? © Сазонов В.Ф., 2012. © kineziolog.bodhy.ru, 2012.
Спинной мозг имеет форму длинного тяжа и располагается вдоль позвоночника.
 На поперечном срезе он похож на бабочку. Только «крылья» этой бабочки называются «рогами» спинного мозга, и их насчитывается 3 пары: передние («моторные»), задние («сенсорные») и маленькие боковые («вегетативные»). Вот такое получается шестирогое чудо природы.
На поперечном срезе он похож на бабочку. Только «крылья» этой бабочки называются «рогами» спинного мозга, и их насчитывается 3 пары: передние («моторные»), задние («сенсорные») и маленькие боковые («вегетативные»). Вот такое получается шестирогое чудо природы. 
Его длина 42-45 см, а далее идёт уже не сам мозг, а «конский хвост» — пучок нервов. На это обязательно надо обратить внимание: на то, что спинной мозг оканчивается где-то на уровне поясницы. Получается странная вещь: поясничный, крестцовый и копчиковый отделы спинного мозга расположены вовсе не в пояснице, крестце и копчике, а гораздо выше!
На фотографии девушки, пытающейся продемонстрировать свой спинной мозг, нижний край сумочки, висящей через плечо, как раз обозначает нижний край спинного мозга. А далее вниз идёт позвоночник, но уже без спинного мозга.
Сущность
Спинной мозг имеет сегментарное строение — и это его главная особенность. Т.е. отдельные его участки как бы повторяются и связаны с определенными участками тела. Всего насчитывается 31-33 сегмента.
Другой особенностью спинного мозга является то, что серое вещество (тела нейронов) находятся в его внутренней части, а на поверхности располагается белое вещество (отростки нейронов). В головном же мозге серое вещество, наоборот, лежит на поверхности в виде коры, а белое вещество спрятано внутрь мозга.
Отделы спинного мозга:
-
Шейный.
-
Грудной.
-
Поясничный.
-
Крестцовый.
-
Копчиковый.
Утолщения:
-
Шейное (управляет верхними конечностями).
-
Пояснично-крестцовое (управляет нижними конечностями).
Нейроны спинного мозга
Количество нейронов в спинном мозге — около 13 млн., и из них всего 3% мотонейронов, которые управляют мышцами. 97% нейронов – вставочные. Как видим, «рабочих» нейронов в нервной системе намного меньше, чем «управленцев»!
Вставочные нейроны относятся и к соматической, и к вегетативной нервной системе.
Классификация нейронов:

1. Эфферентные (исходящие, выносящие). Они относятся к соматической НС, называются мотонейроны, передают возбуждение на скелетные мышцы.
В свою очередь, мотонейроны делятся на несколько групп.
Альфа-мотонейроны (α—мотонейроны) — иннервируют волокна скелетной мускулатуры. У них высокая скорость проведения возбуждения: 70-120 м/с.
Альфа-мотонейроны делятся на:
1) альфа-мотонейроны-1 (быстрые) идут к белым мышечным волокнам, их лабильность (максимальный ритм) — 30 импульсов в секунду. На одном альфа-мотонейроне бывает до 20 тысяч синапсов. Они получают возбуждение, приходящее от кожных рецепторов, проприоцепторов и вышележащих отделов ЦНС,
2) альфа-мотонейроны-2 (медленные) инервируют красные мышечные волокна. Лабильность — 10-15 импульсов в секунду.
Соотношение белых и красных мышц определено у человека генетически.
Гамма-мотонейроны (γ—мотонейроны) — инервируют интрафузальные мышечные волокна мышечных рецепторов. Гамма-мотонейроны изменяют активность мышечных рецепторов. Сами мышечные рецепторы через афферентные и вставочные нейроны посылают импульсы на альфа-мотонейроны. Они активируют собственные альфа-мотонейроны и тормозят мотонейроны мышц-антагонистов. Лабильность (частота импульсации) высокая — 200 импульсов в секунду, но зато небольшая скорость проведения возбуждения — 10-40 м/с.
 2. Афферентные (приносящие, чувствительные).
2. Афферентные (приносящие, чувствительные).
Относятся к соматической НС.
Важно заметить, что их тела расположены не в спинном мозге, а в спинальных ганглиях и ганглиях черепных нервов.
Эти афферентные нейроны проводят возбуждение от рецепторов мышц, сухожилий и кожи. Оканчиваются на вставочных нейронах или, реже, непосредственно на альфа-мотонейронах.
3а. Вставочные соматические нейроны.
Обеспечивают связь между чувствительными афферентными нейронами и эфферентными мотонейронами спинного мозга. Также вставочные нейроны связывают нейроны спинного мозга с ядрами ствола мозга а через них с корой больших полушарий. Лабильность у них высокая — 1000 имп/сек.
3б. Вставочые нейроны вегетативной НС.
Нервные центры симпатической ВНС располагаются в боковых рогах грудного, поясничного и шейного отделов спинного мозга.
Нервные центры парасимпатической ВНС располагаются в крестцовом отделе спинного мозга.
3в. Вставочные ассоциативные нейроны.
Это внутренний аппарат спинного мозга, обеспечивающий связь между его сегментами, а также внутри сегментов. Ассоциативные нейроны участвуют в координации позы, тонуса мышц и движений.
3г. Вставочные нейроны ретикулярной формации спинного мозга.
Они обеспечивают поддержание тонуса самого спинного мозга. Образуют тонкие перекладины серого вещества, идущие в разных направлениях. Сосредоточены в шейных и верхнегрудных сегментах.
Простенькая презентация безусловного рефлекса