- Авторы
- Файлы
Чеснокова Н.П
1
Брилль Г.Е.
1
Моррисон В.В.
1
Понукалина Е.В.
1
Полутова Н.В.
1
1 ФГБОУ ВО «Саратовский Государственный медицинский университет им. В.И. Разумовского Минздрава России»
3.1 Нервная регуляция дыхания
Дыхательный центр представляет собой совокупность нейронов продолговатого мозга, обладающих ритмической активностью и определяющих ритм дыхательных движений. Бульбарный дыхательный центр выполняет две основные функции:
1) регуляцию двигательной активности дыхательных мышц (двигательная функция);
2) гомеостатическую, связанную с изменением характера дыхания при сдвигах газового состава и кислотно-основного равновесия в крови и тканях.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна (длительности вдоха, выдоха, величины дыхательного объема).
Нейроны дыхательного центра расположены в дорсомедиальной и вентролатеральной областях продолговатого мозга, образуя так называемую дорсальную и вентральную дыхательные группы. В указанных дыхательных группах расположены следующие виды нейронов:
1) ранние инспираторные, максимальная частота разряда которых приходится на начало инспирации;
2) поздние инспираторные нейроны, максимальная частота разряда – в конце инспирации;
3) полные инспираторные нейроны, характеризующиеся постоянной активностью в течение фазы вдоха;
4) постинспираторные нейроны, максимальный разряд которых обнаруживается в течение выдоха;
5)экспираторные нейроны, активность которых возрастает во второй части выдоха;
6) преинспираторные нейроны, максимальный пик активности проявляют перед началом вдоха.
В структурах бульбарного дыхательного центра различают так называемые респираторно-связанные нейроны, активность которых совпадает с ритмом дыхания, но они не иннервируют дыхательные мышцы, а обеспечивают иннервацию верхних дыхательных путей.
В соответствии с локализацией нейронов бульбарного дыхательного центра, различают дорсальную дыхательную группу (ДДГ) и вентральную дыхательную группу (ВДГ). Нейроны дорсальной дыхательной группы получают афферентные сигналы от легочных рецепторов растяжения по волокнам n. Vagus. Только часть инспираторных нейронов дорсальной группы дыхательного центра связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контрлатеральной стороной.
Вентральная дыхательная группа расположена латеральнее обоюдного ядра продолговатого мозга, подразделяется на ростральную и каудальную части. Причем, ростральная часть вентральной дыхательной группы представлена ранними, поздними, полными инспираторными и постинспираторными нейронами.
Дорсальная и вентральная группы нейронов в правой и левой половинах продолговотого мозга взаимосвязаны как в пределах одной половины, так и с нейронами противоположной стороны. В синхронизации деятельности контрлатеральных нейронов бульбарного дыхательного центра участвуют проприобульбарные нейроны и экспираторные нейроны комплекса Бетцингера.
Касаясь функциональных особенностей отдельных нейронов бульбарного дыхательного центра, следует отметить, что ранние инспираторные нейроны (активируются в момент вдоха) называют еще проприобульбарными, так как не направляют свои аксоны за пределы дыхательного центра продолговатого мозга и контактируют только с другими типами дыхательных нейронов. Часть полных и поздних инспираторных нейронов направляет свои аксоны к дыхательным мотонейронам спинного мозга. Все экспираторные нейроны каудальной части вентральной дыхательной группы направляют аксоны в спинной мозг. При этом 40% экспираторных нейронов иннервируют внутренние межреберные мышцы, а 60% — мышцы брюшной стенки.
Таким образом, нейроны бульбарного дыхательного центра в зависимости от их значимости в регуляции внешнего дыхания разделяют на три группы:
1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях;
2) нейроны, синаптически связанные с мотонейронами спинного мозга и регулирующие активность мышц вдоха и выдоха;
3) проприобульбарные нейроны, участвующие в генерации дыхательного ритма, аксоны которых обеспечивают связь только с нейронами продолговатого мозга.
Подобно многим физиологическим системам контроля, система управления дыханием организована как контур отрицательной обратной связи.
Афферентация с различных рецепторных зон интегрируется в бульбарном дыхательном центре. Последний, в свою очередь, генерирует импульсацию к мотонейронам спинального отдела дыхательного центра, регулирующего сократительную активность дыхательной мускулатуры.
Важная роль в регуляции внешнего дыхания отводится центрам варолиева моста, в частности, пневмотаксическому центру. Последний включает медиальное, парабрахиальное ядро и ядро Келликера. В парабрахиальном ядре находятся преимущественно инспираторные, экспираторные и фазопереходные нейроны. Ядро Келликера содержит инспираторные нейроны.
Дыхательные нейроны моста участвуют в механизмах смены фаз дыхания, регулируют величину дыхательного объема.
Непосредственными регуляторами сократительной способности дыхательных мышц являются спинальные мотонейроны, получающие информацию по нисходящим ретикулоспинальным путям от бульбарного дыхательного центра.
Как известно, нейроны диафрагмального нерва расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Подавляющее количество волокон диафрагмального нерва являются аксонами α-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен диафрагмы, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны, иннервирующие межреберные мышцы, расположены в передних рогах спинного мозга на уровне TIV-TX, из них часть нейронов регулирует сокращения межреберных мышц, а другая часть – их позно-тоническую активность.
Обращает на себя внимание тот факт, что активность спинальных мотонейронов, обеспечивающих регуляцию двигательной активности межреберных мышц и диафрагмы, в свою очередь, находится под контролем инспираторных нейронов спинного мозга, расположенных на уровне СI-CII вблизи латерального края промежуточной зоны серого вещества.
В обеспечении дыхания, особенно в условиях патологии, участвуют мышцы брюшной стенки, получающие иннервацию от мотонейронов спинного мозга на уровне TIV-LIII.
Двум фазам внешнего дыхания (вдоху и выдоху) соответствуют три фазы активности бульбарного дыхательного центра: инспирация, пассивная контролируемая экспирация и активная экспирация. Во время фазы инспирации диафрагма и наружные межреберные мышцы увеличивают силу сокращения, активируются мышцы гортани, расширяется голосовая щель, снижается сопротивление потоку воздуха. В постинспираторную фазу дыхания происходит медленное расслабление диафрагмы, сокращение мышц гортани, выход воздуха в окружающую среду.
В фазе экспирации – экспираторный поток усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки.
Рефлекторная регуляция дыхания обеспечивается за счет афферентной импульсации в бульбарный дыхательный центр с различных рецепторных зон. Мощной рефлексогенной зоной является слизистая оболочка полости носа, где расположены различные типы механорецепторов, в том числе ирритантные, растяжения, а также болевой чувствительности, обоняния.
Возбуждение этих рецепторов возникает в момент каждого вдоха и приводит к формированию потока афферентной импульсации в ретикулярную формацию ствола мозга с последущей активацией бульбарного дыхательного центра, сосудодвигательного центра, гипоталамических и корковых структур мозга.
Раздражение ирритантных рецепторов слизистой оболочки носа приводит к рефлекторному сужению бронхов, голосовой щели, остановке дыхания на выдохе, развитию брадикардии, а в ряде случаев прекращению сердечных сокращений и другим изменениям (тормозной тригемино-вагусный рефлекс Кречмера ).
Слизистая трахеи и бронхов является слабой рефлексогенной зоной. В стенке крупных внелегочных бронхов и трахеи имеются высокопороговые, низкочувствительные медленноадаптирующиеся, быстроадаптирующиеся и промежуточные механорецепторы, в норме их роль в регуляции дыхания минимальна.
Чувствительность этих рецепторов возрастает при развитии воспалительного процесса в бронхолегочной системе инфекционной или аллергической природы, когда освобождаются медиаторы воспаления и аллергии: гистамин, кинины, лейкотриены, простагландины и др.. Возбудимость рецепторов трахеи и бронхов возрастает и в случае застойных явлений в малом кругу кровообращения, когда прежние объемы воздуха сильно растягивают стенки воздухоносных путей. Афферентация с рецепторов трахеи и бронхов направляется в бульбарный дыхательный центр по чувствительным волокнам n. Vagus, модулируя глубину и частоту дыхательных движений.
Мощной рефлексогенной зоной является паренхима легких, обеспечивающая не только альвеолярное дыхание, но и рефлекторную регуляцию внешнего дыхания.
Основные типы легочных вагусных афферентов включают: медленноадаптирующиеся рецепторы растяжения альвеол, быстроадаптирующиеся рецепторы, С-волокна.
Многочисленные быстроадаптирующиеся рецепторы (БАР) находятся в эпителии внутрилегочных бронхов и бронхиол. Эти рецепторы наиболее чувствительны к следующим типам раздражителей: ирритантным воздействиям, повреждению паренхимы и механическому раздражению дыхательных путей. Возбуждение БАР возникает также при глубоком дыхании, легочной эмболии и капиллярной гипертензии. Афферентация с этих рецепторов распространяется по чувствительным маломиелинизированным волокнам n. Vagus в ретикулярную формацию ствола мозга и бульбарный дыхательный центр, вызывая бронхоконстрикцию, тахипноэ, развитие кашля и тахикардии. Возбуждение этих рецепторов может быть клинически значимым в патогенезе бронхиальной астмы и нарушениях реактивности дыхательных путей.
По данным ряда авторов в паренхиме легких выделяют и БАР рецепторы спадения, реагирующие на спадение альвеол под воздействием внутрилегочных и внелегочных факторов. Афферентация с этих рецепторов поступает в бульбарный дыхательный центр по маломиелинизированным волокнам n. Vagus и обеспечивает развитие тахипноэ.
Медленноадаптирующиеся рецепторы растяжения – важная группа механорецепторов c вагусной афферентацией, расположенных в гладких мышцах воздухоносных путей. Частота импульсов с этих рецепторов возрастает по мере растяжения альвеол вдыхаемым воздухом и распространяется по толстым миелинизированным α-волокнам n. Vagus в бульбарный дыхательный отдел, обеспечивая формирование рефлекса Геринга-Брейера. Последний контролирует частоту и глубину дыхания, имеет физиологическое значение при дыхательных объемах превышающих 1 л (у взрослых при физической нагрузке). Рефлекс Геринга-Брейера более важен для регуляции дыхательного акта у новорожденных, а также в условиях патологии как один из механизмов реализации инспираторной, экспираторной и смешанной одышек.
Третьей группой легочных механорецепторов являются С-волокна – тонкие миелинизированные вагусные афференты. С – волокна оканчиваются в паренхиме легких, в бронхах и кровеносных сосудах, активируются экзогенными раздражителями и медиаторами альтерации. Активация С-волокон приводит к тахипноэ, брадикардии, гиперсекреции слизи. В состав С-волокон входят J-рецепторы, расположенные в альвеолярных перегородках в контакте с капиллярами (юкстакапиллярные рецепторы), чувствительные к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам. Активация J-рецепторов вызывает закрытие гортани и апноэ, за которыми следует частое поверхностное дыхание, гипотензия и брадикардия.
Важная роль в рефлекторной регуляции дыхания отводится проприорецепторам суставов грудной клетки, межреберных мышц, диафрагмы, сухожильным рецепторам. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которая через α-мотонейроны повышает активность α-мотонейронов и дозирует таким образом мышечное усилие.
В регуляции активности бульбарного дыхательного центра и внешнего дыхания принимает участие и афферентация с висцеральных рецепторов и рецепторов кожи, о чем свидетельствует развитие гипервентиляции легких при болевом и термическом раздражении.
3.2. Механизмы гуморальной регуляции дыхания
Важная роль в регуляции дыхания отводится хеморецепторам.
Изменения газового состава крови (РаО2, РаСО2) влияют на активность дыхательного центра путем возбуждения хеморецепторов каротидных и аортальных телец (периферические рецепторы), а также хеморецепторов вентральной зоны продолговатого мозга и дорсального дыхательного ядра (центральные рецепторы). Периферические хеморецепторы (рис.5) обеспечивают регуляцию частоты дыхательных движений. Адекватным раздражителем для них является уменьшение РО2 артериальной крови, в меньшей степени – увеличение РСО2 и снижение рН. Периферические хеморецепторы расположены у бифуркации общих сонных артерий на внутреннюю и наружнюю. Несмотря на свой миниатюрный размер, каротидные тельца интенсивно кровоснабжаются (1,4-2 л/мин на 100 г ткани). Этот орган особенно чувствителен к колебаниям кислорода в артериальной крови. При Ра О2 в пределах 60-80 мм рт. ст. наблюдается слабое усиление вентиляции, при Ра О2 ниже 50 мм рт. ст. возникает выраженная гипервентиляция легких. Ра СО2 и рН крови потенцируют эффекты гипоксемии на артериальные хеморецепторы и не являются адекватными раздражителями для этих рецепторов. После двустороннего удаления каротидных телец гипоксический вентиляторный ответ у человека исчезает. При отсутствии хеморецепторной стимуляции, например, при глубокой гипокапнии, повреждении синокаротидной зоны (опухоли, коллагенозы, травмы) ритмогенез дыхания снижается и полностью прекращается.
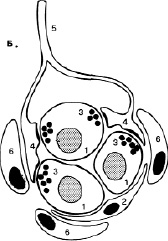
Рис. 5. Каротидное тельце: 1-хеморецепторные клетки; 2-поддерживающие клетки; 3-синаптические пузырьки; 4-чувствительные нервные окончания; 5-нервное волокно
Центральные хемочувствительные клетки реагируют на отклонения РСО2 и [H+] во внеклеточной жидкости внутримозгового интерстициального пространства, регулируют глубину вдоха. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Одной из причин высокой скорости вентиляторного ответа на гиперкапнию является легкость диффузии СО2 через барьерную систему кровь-головной мозг. Более того, повышенное РСО2 вызывает расширение сосудов, особенно церебральных, способствуя тем самым усилению диффузии СО2 через гемато-энцефалический барьер.
Библиографическая ссылка
Чеснокова Н.П, Брилль Г.Е., Моррисон В.В., Понукалина Е.В., Полутова Н.В. ЛЕКЦИЯ 3 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ ДЫХАНИЯ // Научное обозрение. Медицинские науки. – 2017. – № 2.
– С. 36-39;
URL: https://science-medicine.ru/ru/article/view?id=972 (дата обращения: 03.04.2023).
Предлагаем вашему вниманию журналы, издающиеся в издательстве «Академия Естествознания»
(Высокий импакт-фактор РИНЦ, тематика журналов охватывает все научные направления)
Диафрагма (англ. diaphragm) – необычная мышца, так как она представляет собой несколько круглый мышечный лист с центральным сухожилием и костным прикреплением только по её окружности. Во время сокращения он тянется от периферии к центральному сухожилию.

Поскольку действия диафрагмы увеличивают объем грудной клетки, диафрагма, несомненно, является мышцей вдоха. Диафрагма сжимается от своих периферийных креплений на нижних ребрах и позвонках и натягивает центральное сухожилие, тем самым оттягивая центральное сухожилие ниже и увеличивая вертикальную длину грудной полости.
По мере опускания грудного дна объем брюшной полости уменьшается, а брюшное давление увеличивается. Если брюшная стенка остается расслабленной, внутренние органы брюшной полости выдвигаются вперед, и переднезадний диаметр брюшной полости увеличивается. Хотя диафрагма расположена глубже нижних ребер и не может быть пропальпирована, её сокращение легко определить, наблюдая за движениями содержимого брюшной полости.
Внутренние органы брюшной полости ограничивают полное опускание диафрагмы, допустимое сократительной длиной мышечных волокон диафрагмы. Продолжающееся сокращение диафрагмы после того, как она достигает максимального спуска на внутренние органы, поднимает нижние ребра, продолжая увеличивать объем грудной клетки.
Начало
Диафрагма прикрепляется по периферии в трех частях: грудной, реберной и поясничной. Грудинная часть прикрепляется к задней поверхности мечевидного отростка. Это прикрепление может отсутствовать. Реберная часть прикрепляется к глубоким поверхностям нижних шести реберных хрящей и ребер. Поясничная часть прикрепляется к поясничным позвонкам и двум апоневротическим дугам, а также медиальной и боковой дугообразным связкам.
Прикрепление
Периферийные прикрепления сходятся, чтобы прикрепиться к центральному сухожилию, которое не имеет костных прикреплений.
Иннервация
Диафрагмальный нерв (С3-5).
Кровоснабжение
Подреберные и нижние пять межреберные артерии, нижние диафрагмальные артерии, верхние диафрагмальные артерии.
Функция
Диафрагма является одной из основных дыхательных мышц. Когда мышечные волокна сокращаются, диафрагма уплощается. Это увеличивает объем грудной полости по вертикали, что снижает внутрилегочное давление, и воздух поступает в легкие. Когда диафрагма расслабляется, объем грудной клетки уменьшается, внутрилегочное давление повышается, и воздух выходит из легких.
Когда диафрагма работает с переднелатеральными мышцами живота, сокращение диафрагмы способствует повышению внутрибрюшного давления. Это необходимо при таких действиях, как рвота, дефекация, мочеиспускание и роды. Другая функция диафрагмы заключается в обеспечении прохода определенных структур из грудной клетки в брюшную полость (нижняя полая вена, пищевод и аорта).
Пальпация
Пальпация невозможна.
Клиническая значимость
Слабость и / или паралич диафрагмы могут возникать у лиц с травмой верхнего шейного отдела спинного мозга (С3), которые также проявляются слабостью или утратой функций межреберных и лестничных мышц. Такие пациенты имеют глубокое нарушение вдоха и нуждаются, по крайней мере, в периодической механической вентиляции легких. Слабость диафрагмы, даже при неповрежденных мышцах грудной клетки, приводит к существенному нарушению работы инспираторного аппарата. Изолированная слабость диафрагмы создает картину парадоксального дыхания, при которой окружность брюшной полости уменьшается во время вдоха.
Икота
Очень распространенным нарушением работы диафрагмы, которое в какой-то момент поражает большинство людей, является икота. Икота возникает из-за непроизвольного, прерывистого сокращения мышц. Обычно они вызваны потреблением больших объемов пищи в течение короткого промежутка времени.
Грыжи
Грыжи могут возникать через диафрагму. На уровне пищеводного отверстия диафрагмы желудок может образовывать грыжу в заднем средостении, состояние, известное как грыжа пищеводного отверстия диафрагмы. Диафрагмальная грыжа может быть врожденной. Они возникают в результате аномалий развития диафрагмы у плода. Органы брюшной полости могут выступать в грудную полость и нарушать развитие легких, вызывая проблемы в развитии легких и функционировании легких после рождения. Диафрагмальные грыжи также могут быть приобретенными. Обычно это происходит в результате травмы тупым предметом, например, в результате дорожно-транспортного происшествия или сильного падения. Для устранения диафрагмальной грыжи требуется хирургическое вмешательство.
Упражнения
см. упражнения для диафрагмы
Стретчинг
см. растяжка диафрагмы
Массаж
см. массаж диафрагмы
Миофасциальный релиз
см. МФР диафрагмы
Диафрагма,
м.
френикус
– это
подвижная мышечно-сухожильная перегородка
между грудной и брюшной полостями.
Имеет куполообразную форму, выпуклая
часть диафрагмы обращена в грудную
полость, вогнутая – в брюшную. Правый
купол (5-е межреберье) выше левого
обусловлен расположением под ним печени.
Иннервируется
диафрагмальным нервом из шейного
сплетения. Функция.
Диафрагма – это
главная дыхательная мышца и орган
брюшного пресса.
При сокращении диафрагмы ее купол
уплощается, и тогда грудная полость
увеличивается, а брюшная уменьшается.
При одновременном сокращении с мышцами
живота диафрагма повышает внутрибрюшное
давление. Диафрагма имеет
3 части:
грудинную,
реберную и поясничную, от
которых начинаются
мышечные пучки,
они
сходятся
в сухожильном
центре диафрагмы. Поясничная часть
начинается на поясничных позвонках
правой
и левой ножками
и на двух
дугообразных связках медиальной и
латеральной. Ножки
диафрагмы,
сближаясь ограничивают
аортальное
отверстие,
затем расходятся и снова сходятся,
ограничивая пищеводное
отверстие.
Через отверстие
в сухожильном центре
диафрагмы проходит нижняя
полая вена.
Через
мышечные пучки диафрагмы
проходят непарная
вена
(справа)
и полунепарная (слева),
правый и левый симпатические
стволы,
большой и малый внутренностные
нервы.
Реберная
часть
начинается мышечными пучками от
6-7 нижних ребер,
грудинная
часть,
самая узкая от задней
поверхности грудины,
обе части тоже сходятся своими пучками
в сухожильном центре. Между
частями
диафрагмы имеются участки,
лишенные мышечных пучков,
где есть только
брюшина и плевра
и брюшная
и грудная фасции. Это
пояснично-реберные
и грудино-реберные треугольники,
слабые
места
диафрагмы,
места выхода диафрагмальных грыж.
30. Мышцы живота, иннервация.
Различают
мышцы боковой стенки живота, передней
стенки и задней стенки живота.
Боковую
стенку живота
образуют 3
мышцы,
которые располагаются
послойно:
наружная
косая, внутренняя косая и поперечная
мышцы живота. Их
волокна перекрывают друг друга под
углом в 90 градусов.
Наружная
косая м. живота (м.обликвус
экстернус абдоминис) –
ее иннервирует подвздошно-паховый
и подвздошно-подчревный
нервы
(из поясничного сплетения), и нижние
межреберные нервы.
Внутренняя
косая
м. живота и поперечная
м.
живота имеют ту
же
иннервацию.
Эти
мышцы начинаются от нижних шести ребер
(наружная косая), от подвздошного гребня
и паховой связки (внутренняя косая) и
поперечных отростков поясничных
позвонков и нижних ребер (поперечная).
Перейдя
на переднюю стенку все три мышцы переходят
в апоневрозы
(сухожильные растяжения). Апоневрозы
с правой и левой стороны идут навстречу
друг другу и вплетаются посредине
живота, образуя
белую
линию живота. Апоневроз наружной косой
м. живота своим нижним краем натягивается
между передней верхней подвздошной
остью и лобковым симфизом и образует
паховую связку
(пупартову связку). У
места прикрепления к симфизу апоневроз
делится на две ножки: медиальную – к
лобковому симфизу и латеральную (к
лобковому бугорку).
Поперечная
м. живота
переходит
в апоневроз у латерального края прямой
мышцы живота, образуя
полулунную
линию (спигелева линия).
Мышцы
передней стенки.
Прямая
м. живота (м. ректус абдоминис) –
лежит по бокам от срединной линии живота,
состоит из нескольких брюшков, разделенных
3-4 сухожильными перемычками. Она
начинается от лобкового гребня и симфиза,
идет вверх и прикрепляется к мечевидному
отростку грудины и хрящам 5-7 ребер.
Пирамидальная
м.,
небольшая идет от лобкового симфиза и
вплетается в белую линию живота. Эти
мышцы иннерв.
подвздошно-подчревным нервом и нижними
межреберными нервами.
На
задней
стенке живота
находится квадратная
мышца поясницы
(м. квадратус люмборум), она иннервир.
мышечными
ветвями поясничного сплетения.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
From Wikipedia, the free encyclopedia
«Phrenectomy» redirects here. Not to be confused with Frenectomy.
| Phrenic nerve | |
|---|---|

The phrenic nerve as it passes through the thorax to supply the diaphragm. |
|

The phrenic nerve emerges from the cervical plexus, with the right brachial plexus shown here. |
|
| Details | |
| From | C3–C5 of cervical plexus |
| Innervates | Diaphragm |
| Identifiers | |
| Latin | nervus phrenicus |
| MeSH | D010791 |
| TA98 | A14.2.02.028 |
| TA2 | 6380 |
| FMA | 6191 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
The phrenic nerve is a mixed motor/sensory nerve that originates from the C3-C5 spinal nerves in the neck. The nerve is important for breathing because it provides exclusive motor control of the diaphragm, the primary muscle of respiration. In humans, the right and left phrenic nerves are primarily supplied by the C4 spinal nerve, but there is also a contribution from the C3 and C5 spinal nerves. From its origin in the neck, the nerve travels downward into the chest to pass between the heart and lungs towards the diaphragm.
In addition to motor fibers, the phrenic nerve contains sensory fibers, which receive input from the central tendon of the diaphragm and the mediastinal pleura, as well as some sympathetic nerve fibers. Although the nerve receives contributions from nerve roots of the cervical plexus and the brachial plexus, it is usually considered separate from either plexus.
The name of the nerve comes from Ancient Greek phren ‘diaphragm’.[1]
Structure[edit]
The phrenic nerve originates in the phrenic motor nucleus in the ventral horn of the cervical spinal cord. It descends obliquely with the internal jugular vein across the anterior scalene, deep to the prevertebral layer of deep cervical fascia and the transverse cervical and suprascapular arteries. On the left, the phrenic nerve crosses anterior to the first part of the subclavian artery. On the right, it lies on the anterior scalene muscle and crosses anterior to the 2nd part of the subclavian artery. On both sides, the phrenic nerve usually runs posterior to the subclavian vein as it enters the thorax where it runs anterior to the root of the lung and between the fibrous pericardium and mediastinal parietal pleura.[2]
- The right phrenic nerve passes over the brachiocephalic artery, posterior to the subclavian vein, and then crosses the root of the right lung anteriorly and then leaves the thorax by passing through the vena cava hiatus opening in the diaphragm at the level of T8. The right phrenic nerve passes over the right atrium.
- The left phrenic nerve passes over the pericardium of the left ventricle and pierces the diaphragm separately.
The pericardiacophrenic arteries and veins travel with their respective phrenic nerves.
The phrenic nerve can be marked by a line connecting these two points:
- 1st point can be labelled 3.5 cm at the level of the thyroid cartilage from the midsagittal plane.
- 2nd point is at the medial end of the clavicle.
Variation[edit]
As with most nerves in the neck, multiple anatomic variants have been described. Notably, there may be variability in the course of the phrenic nerve in the retro-clavicular region such that the nerve courses anterior to the subclavian vein, rather than its typical position posterior to the vein (between the subclavian vein and artery).[3] This variant may predispose the phrenic nerve to injury during subclavian vascular cannulation.
In addition, an accessory phrenic nerve is commonly identified, present in up to 75% of a cadaveric study.[3]
In canines, the phrenic nerve arises from C5-C7 with occasional small contributions from C4.[4] In the cat, horse, ox, and small ruminant the phrenic nerve arises variably from C4-C7.
Function[edit]
Both of these nerves supply motor fibers to the diaphragm and sensory fibers to the fibrous pericardium, mediastinal pleura, and diaphragmatic peritoneum.
Some sources describe the right phrenic nerve as innervating the gallbladder, other sources make no such mention.[5]
The right phrenic nerve may also supply the capsule of the liver.[6]
Clinical significance[edit]

Left phrenic nerve palsy (right image side) in fluoroscopy: forced inspiration with the closed mouth leads to paradox elevation of the paralytic left diaphragm while the healthy right side moves down.
Pain arising from structures supplied by the phrenic nerve is often «referred» to other somatic regions served by spinal nerves C3-C5. For example, a subphrenic abscess beneath the right diaphragm might cause a patient to feel pain in the right shoulder.
Irritation of the phrenic nerve (or the tissues it supplies) leads to the hiccup reflex. A hiccup is a spasmodic contraction of the diaphragm, which pulls air against the closed folds of the larynx.
The phrenic nerve must be identified during thoracic surgery and preserved. To confirm the identity of the phrenic nerve, a doctor may gently manipulate it to elicit a dartle (diaphragmatic startle) response.[7] The right phrenic nerve may be crushed by the vena cava clamp during liver transplantation.[8] Severing the phrenic nerve, or a phrenectomy,[9] will paralyse that half of the diaphragm. Bilateral diaphragmatic paralysis or BDP can also be caused by spinal cord injury, motor neuron disease, infection, pneumonia, sarcoidosis, multiple sclerosis, polyneuropathy, myopathy and amyotrophy, cardiac surgery, lung transplantation, or mediastinal tumors.[10][11] Diaphragm paralysis is best demonstrated by sonography.[8] Breathing will be made more difficult but will continue provided the other nerve is intact.
The phrenic nerve arises from the neck (C3-C5) and innervates the diaphragm, which is much lower. Hence, patients suffering spinal cord injuries below the neck are still able to breathe effectively, despite any paralysis of the lower limbs.
Brachial plexus injuries can cause paralysis in various regions in the arm, forearm, and hand depending on the severed nerves. The resulting palsy has been clinically treated using the phrenic nerve as a donor for neurotization of the musculocutaneous nerve and the median nerve.[12] This treatment has a high success rate (84.6%) in partial to full restoration of the innervation to the damaged nerve.[12] Furthermore, this procedure has resulted in restored function to nerves in the brachial plexus with minimal impact to respiratory function of the phrenic nerve. The instances where pulmonary vital capacity is reduced have typically been a result of use of the right phrenic as the donor for the neurotization whereas use of left phrenic nerve has not been significantly linked to reduced pulmonary vital capacity.[13]
See also[edit]
References[edit]
- ^ O’Rahilly, Ronan (2008). Basic Human Anatomy. Hanover, New Hampshire: Geisel School of Medicine. Retrieved 3 April 2019.
- ^ Moore, Keith L. (1999). Clinically oriented anatomy. Philadelphia: Lippincott Williams & Wilkins. ISBN 978-0-683-06141-3.[page needed]
- ^ a b Prakash; Prabhu, L. V.; Madhyastha, S; Singh, G (2007). «A variation of the phrenic nerve: Case report and review» (PDF). Singapore Medical Journal. 48 (12): 1156–7. PMID 18043847.
- ^ Evans, Howard (1979). Miller’s Anatomy of the Dog. Philadelphia, PA: W. B. Saunders Company. p. 978. ISBN 978-0-7216-3438-8.
- ^ Alexander, William (1940). «The innervation of the biliary system». Journal of Comparative Neurology. 72 (2): 357–370. doi:10.1002/cne.900720205. S2CID 83974380.
- ^ Lautt, W. Wayne (2009). Hepatic Nerves. Morgan & Claypool Life Sciences.
- ^ Dalman, Ronald; Thompson, Robert (2015). «7. Neurogenic Thoracic Outlet Syndrome Exposure and Decompression: Supraclavicular». Operative Techniques in Vascular Surgery. Philadelphia, PA: Wolters Kluwer Health. p. 50. ISBN 9781451190205.
- ^ a b McAlister, Vivian C.; Grant, David R.; Roy, Andre; Brown, William F.; Hutton, Linda C.; Leasa, David J.; Ghent, Cameron N.; Veitch, James E.; Wall, William J. (1993). «Right phrenic nerve injury in orthotopic liver transplantation». Transplantation. 55 (4): 826–30. doi:10.1097/00007890-199304000-00027. PMID 8475559.
- ^ Hine, Maynard Kiplinger (1975). Review of dentistry: questions and answers (6th ed.). Mosby. p. 421. ISBN 978-0-8016-2196-3.
- ^ Mizubuti GB, Wang L, Ho AM, Tanzola RC, Leitch J (November 2017). «Perioperative Management for Abdominal Surgery in Bilateral Diaphragmatic Paralysis: A Case Report and Literature Review». A & A Case Rep. 9 (10): 280–282. doi:10.1213/XAA.0000000000000592. PMID 28691979. S2CID 44462495.
- ^ Chen HY, Chen HC, Lin MC, Liaw MY (August 2015). «Bilateral Diaphragmatic Paralysis in a Patient With Critical Illness Polyneuropathy: A Case Report». Medicine (Baltimore). 94 (31): e1288. doi:10.1097/MD.0000000000001288. PMC 4616567. PMID 26252301.
- ^ a b Gu YD, Wu MM, Zhen YL, Zhao JA, Zhang GM, Chen DS, Yan JG, Cheng XM (1989). «Phrenic nerve transfer for brachial plexus motor neurotization». Microsurgery. 10 (4): 287–9. doi:10.1002/micr.1920100407. PMID 2593799. S2CID 74732561.
- ^ Luedemann W, Hamm M, Blömer U, Samii M, Tatagiba M (March 2002). «Brachial plexus neurotization with donor phrenic nerves and its effect on pulmonary function». J Neurosurg. 96 (3): 523–6. doi:10.3171/jns.2002.96.3.0523. PMID 11883837.
External links[edit]
- Anatomy figure: 19:04-05 at Human Anatomy Online, SUNY Downstate Medical Center — «Left side of the mediastinum.»
- Anatomy figure: 25:03-15 at Human Anatomy Online, SUNY Downstate Medical Center — «Diagram of the cervical plexus.»