| Кохлеарные ядра | |
|---|---|

Рассечение мозговой ствол. Вид сверху. («Кохлеарное ядро» указано слева, пятое снизу.) |
|

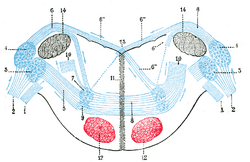
Терминальные ядра улиткового нерва с их верхними связями. (Схема.) Вестибулярный нерв с его конечными ядрами и их эфферентными волокнами подавлен. С другой стороны, чтобы не закрывать трапециевидное тело, эфферентные волокна терминальных ядер с правой стороны были резецированы на значительной части их протяженности. Таким образом, трапециевидное тело показывает только половину волокон, а именно те, которые идут слева.
|
|
| Подробности | |
| Часть | мозговой ствол |
| Система | Слуховая система |
| Артерия | AICA |
| Идентификаторы | |
| латинский | ядра улитки |
| MeSH | D017626 |
| NeuroNames | 720 |
| НейроЛекс Я БЫ | birnlex_1151 |
| TA98 | A14.1.04.247 A14.1.05.430 |
| TA2 | 6006, 6007 |
| FMA | 72240 |
| Анатомические термины нейроанатомии
[редактировать в Викиданных ] |
В кохлеарный ядерный (CN) комплекс состоит из двух ядра черепных нервов в человеке мозговой ствол, то вентральное кохлеарное ядро (VCN) и дорсальное кохлеарное ядро (DCN). Вентральное ядро улитки не слоистое, тогда как дорсальное ядро улитки слоистое. Волокна слухового нерва, волокна, которые проходят через слуховой нерв (также известный как кохлеарный нерв или восьмой черепной нерв) несут информацию от внутреннего уха, улитка на той же стороне головы к нервному корешку вентрального ядра улитки. В корешке нерва волокна разветвляются, иннервируя вентральное ядро улитки и глубокий слой дорсального ядра улитки. Таким образом, вся акустическая информация поступает в мозг через ядра улитки, где начинается обработка акустической информации. Выходы из ядер улитки принимаются в более высоких областях слуховой ствол мозга.
Структура
Ядра улитки (CN) расположены на дорсо-латеральной стороне мозговой ствол, охватывающий стык мосты и мозговое вещество.
- В вентральное кохлеарное ядро(VCN) на вентральной части ствола мозга, вентролатерально к нижний цветонос.
- В дорсальное кохлеарное ядро (DCN), также известный как tuberculum acusticum или акустический бугорок, изгибается над VCN и огибает ножку мозжечка.
- VCN далее разделяется нервным корешком на задневентральное ядро кохлеара (PVCN) и антеровентральное ядро кохлеара (AVCN).[1]
Проекции на ядра улитки
Основной вход в кохлеарное ядро поступает от слухового нерва, части VIII черепного нерва ( вестибулокохлеарный нерв ). Волокна слухового нерва образуют высокоорганизованную систему связей в соответствии с их периферической иннервацией улитки. Аксоны из спиральный узел клетки более низких частот иннервируют вентролатеральные части вентрального кохлеарного ядра и латерально-вентральные части дорсального кохлеарного ядра. Аксоны от более высокой частоты кортиевый орган волосковые клетки проецируются на дорсальную часть вентрального ядра улитки и дорсально-медиальные части дорсального ядра улитки. Среднечастотные проекции оказываются между двумя крайностями; таким образом тонотопическая организация, установленная в улитке, сохраняется в ядрах улитки. Эта тонотопная организация сохраняется, потому что только несколько внутренних волосковых клеток синапсируются на дендритах нервной клетки в спиральном ганглии, а аксон от этой нервной клетки синапсируется только на очень небольшом количестве дендритов в ядре улитки. В отличие от VCN, который получает весь акустический вход от слухового нерва, DCN получает входной сигнал не только от слухового нерва, но также получает акустический вход от нейронов в VCN (Т-звездчатых клетках). Таким образом, DCN в некотором смысле является сенсорным ядром второго порядка.
Долгое время считалось, что ядра улитки получают сигнал только от ипсилатерального уха. Однако есть доказательства стимуляции от контралатерального уха через контралатеральный CN,[2] а также соматосенсорные части мозга.[3]
Проекции ядер улитки
Есть три основных пучка волокон, аксоны ядерных нейронов улитки, которые переносят информацию от ядер улитки к мишеням, которые в основном находятся на противоположной стороне мозга. Сквозь мозговое вещество, одна проекция идет на контралатеральную высший оливарный комплекс (SOC) через трапециевидное тело, в то время как другая половина стреляет в ипсилатеральный SOC. Этот путь называется вентральной акустической полосой (ВАШ или, чаще, трапециевидным телом). Другой путь, называемый дорсальной акустической полоской (DAS, также известной как полоса фон Монакова), поднимается над мозговым слоем в мосты где он попадает в ядра боковой лемниск наряду с его родственником, промежуточной акустической полосой (IAS, также известной как полоса Хельда). IAS пересекает продолговатый мозг перед тем, как присоединиться к восходящим волокнам контралатерального латерального лемниска. Боковой лемниск содержит клетки ядер латерального лемниска и, в свою очередь, выступает в нижний бугорок. Нижний бугорок получает прямые моносинаптические проекции от верхнего оливкового комплекса, контралатеральной дорсальной акустической полоски, некоторых классов звездчатых нейронов VCN, а также от различных ядер латерального лемниска.
Большинство этих входов оканчиваются в нижнем бугорке, хотя есть несколько небольших выступов, которые обходят нижний бугорок и проецируются на медиальный колликулюс или другие структуры переднего мозга.
- Медиальная верхняя оливковая (MSO) через трапециевидное тело (TB) — Ипсилатеральная и контралатеральная стимуляция звуков низкой частоты.
- Боковая верхняя олива (LSO) напрямую и через TB — Ипсилатеральная стимуляция высокочастотными звуками.
- Медиальное ядро трапециевидное тело (MNTB) — Контралатеральная стимуляция.
- Нижний холмик — Контралатеральная стимуляция.
- Периоливарные ядра (ПОН) — Ипсилатеральная и контралатеральная стимуляция.
- Боковой лемниск (LL) и лемнисковые ядра (LN) — Ипсилатеральная и контралатеральная стимуляция.
Гистология
Три типа основных клеток передают информацию из вентрального ядра улитки: кустистые клетки, звездчатые клетки и клетки осьминога.
- Кустистые клетки обнаруживаются в основном в переднем вентральном ядре улитки (AVCN). В дальнейшем их можно разделить на большие сферические, мелкие сферические и шаровидные кустистые клетки, в зависимости от их внешнего вида, а также их расположения. Внутри AVCN есть область больших сферических клеток; каудальнее к нему расположены более мелкие сферические клетки, а глобулярные клетки занимают область вокруг нервного корешка. Важное различие между этими подтипами заключается в том, что они проецируются на разные цели в высшем оливковом комплексе. Крупные сферические густые клетки выступают на ипсилатеральную и контралатеральную медиальную верхнюю оливу. Глобулярные кустистые клетки проецируются на контралатеральное медиальное ядро трапециевидного тела, а маленькие сферические кустистые клетки, вероятно, проецируются на латеральную верхнюю оливу. У них есть несколько (1-4) очень коротких дендритов с многочисленными мелкими ветвлениями, которые делают его похожим на «куст». Кустообразные клетки обладают особыми электрическими свойствами, которые позволяют им передавать информацию о времени от слухового нерва к более центральным областям слуховой системы. Поскольку кустистые клетки получают входной сигнал от множества волокон слухового нерва, которые настроены на одинаковые частоты, пушистые клетки могут улучшить точность информации о времени, по существу усредняя дрожание во времени входных сигналов. Кустоватые клетки также могут подавляться звуками, близкими к частоте, на которую они настроены, что приводит к еще более резкой настройке, чем это наблюдается в волокнах слухового нерва. Эти клетки обычно иннервируются только несколькими волокнами слухового нерва, которые доминируют в его возбуждении. Эти афферентные нервные волокна обвивают свои терминальные ветви вокруг всей сомы, создавая большой синапс на густых клетках, называемый «конец лампы Held «. Следовательно, единичная запись электрически стимулированного кустистого нейрона, как правило, дает ровно один потенциал действия и составляет первичный ответ.
- Звездчатые клетки (также известные как мультиполярные клетки), имеют более длинные дендриты, расположенные параллельно пучкам волокон слухового нерва. Их также называют чопперными клетками в связи с их способностью запускать серию потенциалов действия с регулярным интервалом в течение тонального или шумового стимула. Схема измельчения присуща электрическая возбудимость звездчатой клетки, и частота импульсов больше зависит от силы слухового сигнала, чем от частоты. Каждая звездчатая ячейка имеет узкую настройку и имеет тормозящие боковые полосы, что позволяет населению звездчатых клеток кодировать спектр звуков, усиливая спектральные пики и спады. Эти нейроны обеспечивают акустический ввод в DCN.
- Клетки осьминога находятся в небольшой области заднего вентрального ядра улитки (PVCN). Отличительными чертами этих клеток являются их длинные, толстые дендриты в форме щупалец, которые обычно исходят с одной стороны тела клетки. Клетки осьминога вырабатывают «начальную реакцию» на простые тональные стимулы. То есть они реагируют только на появление широкополосного стимула. Клетки осьминога могут активироваться с высочайшей временной точностью среди нейронов мозга. Электрические стимулы слухового нерва вызывают в клетках осьминога ступенчатый возбуждающий постсинаптический потенциал. Эти EPSP очень краткие. Считается, что клетки осьминога важны для извлечения информации о времени. Сообщалось, что эти клетки могут реагировать на последовательность щелчков с частотой 800 Гц.
Два типа основных ячеек передают информацию из дорсальное кохлеарное ядро (DCN) к контралатеральному нижнему бугорку. Основные ячейки получают две системы входов. Акустический сигнал поступает в глубокий слой несколькими путями. Возбуждающий акустический сигнал поступает от волокон слухового нерва, а также от звездчатых клеток VCN. Акустический вход также передается через тормозящие интернейроны (туберкуловентральные клетки DCN и «широкополосные ингибиторы» в VCN). Через самый внешний молекулярный слой DCN получает другие типы сенсорной информации, в первую очередь информацию о расположении головы и ушей, через параллельные волокна. Эта информация распространяется через мозжечковую цепь, которая также включает тормозящие интернейроны.
- Веретенообразные клетки (также известен как пирамидные клетки). Веретенообразные клетки интегрируют информацию через два пучка дендритов, апикальные дендриты получают мультисенсорную, возбуждающую и тормозную информацию через самый внешний молекулярный слой, а базальные дендриты получают возбуждающий и тормозной акустический вход от базальных дендритов, которые простираются в глубокий слой. Считается, что эти нейроны позволяют млекопитающим анализировать спектральные сигналы, которые позволяют нам локализовать звуки на высоте и когда мы теряем слух на одно ухо.
- Гигантские клетки также интегрируют входные данные из молекулярного и глубокого слоев, но преобладают входные данные из глубокого слоя. Неясно, какова их роль в слушании.
Функция
Ядерный комплекс улитки является первым интегративным, или обрабатывающим, этапом слуховая система. Информация в ядра поступает из ипсилатеральный улитка через кохлеарный нерв. В ядрах улитки выполняется несколько задач. Распределяя акустический вход на несколько типов основных клеток, слуховой путь подразделяется на параллельные восходящие пути, которые могут одновременно извлекать различные типы информации. Клетки вентрального ядра улитки извлекают информацию, которая переносится слуховым нервом во время возбуждения и в структуре активации популяции волокон слухового нерва. Клетки дорсального ядра улитки выполняют нелинейный спектральный анализ и помещают этот спектральный анализ в контекст расположения головы, ушей и плеч и отделяют ожидаемые, самогенерируемые спектральные сигналы от более интересных, неожиданных спектральных сигналов, используя вклад от слуховая кора, понтинные ядра, тройничный узел и ядро, ядра спинного столба а второй ганглий дорзального корня. Вполне вероятно, что эти нейроны помогают млекопитающим использовать спектральные сигналы для ориентации на эти звуки. Информация используется высшими отделами ствола мозга для достижения дальнейших вычислительных целей (таких как определение местоположения источника звука или улучшение соотношение сигнал шум ). Поступающие из этих других областей мозга, вероятно, играют роль в локализации звука.
Чтобы более подробно понять специфические функции ядер улитки, сначала необходимо понять, как звуковая информация представлена волокнами слуховой нерв. Вкратце, в каждом из двух слуховых нервов около 30 000 волокон слухового нерва. Каждое волокно является аксоном спиральный узел ячейка, которая представляет определенную частоту звука и определенный диапазон громкости. Информация в каждом нервном волокне представлена скоростью потенциалы действия а также конкретное время индивидуальных потенциалов действия. Особая физиология и морфология каждого типа клеток ядра улитки усиливает различные аспекты звуковой информации.
Смотрите также
- Слуховая система
- Кохлеарный нерв
- Униполярная щеточная ячейка
Дополнительные изображения
-

Рассечение ствола мозга. Боковой вид.
-

Схематично изображены ядра черепных нервов; вид со спины. Моторные ядра выделены красным цветом; сенсорный синий.
-

Схематично изображены первичные терминальные ядра афферентных (сенсорных) черепных нервов; вид сбоку.
-

Схема, показывающая ход волокон лемниска; медиальный лемниск синий, латеральный красный.
-

Поперечный разрез нижнего моста, показывающий ядро улитки (№1), помеченное вверху справа.





Рекомендации
Эта статья включает текст в всеобщее достояние из стр. 788 20-го издания Анатомия Грея (1918)Молодой ЭД, Спиру Г.А., Райс Дж.Дж., Фойгт Х.Ф. (июнь 1992 г.). «Нейронная организация и ответы на сложные раздражители в дорсальном ядре улитки». Филос. Пер. R. Soc. Лондон. B Biol. Наука. 336 (1278): 407–13. Дои:10.1098 / рстб.1992.0076. PMID 1354382.
- ^ Миддлбрукс, Дж. К. (2009). «Слуховая система: центральные пути». В Сквайре (ред.). Энциклопедия неврологии. Академическая пресса. стр. 745–752, здесь: с. 745 ф.
- ^ Дэвис К.А. (сентябрь 2005 г.). «Контралатеральные эффекты и бинауральные взаимодействия в дорсальном ядре улитки». J. Assoc. Res. Отоларингол. 6 (3): 280–96. Дои:10.1007 / s10162-005-0008-5. ЧВК 2504593. PMID 16075189.
- ^ Шор, С.Е. (2009). «Слуховые / соматосенсорные взаимодействия». В Сквайре (ред.). Энциклопедия неврологии. Академическая пресса. С. 691–5.
внешняя ссылка
- Университет Буффало на Библиотека Конгресса Веб-архивы (архивировано 27 ноября 2001 г.)
- Иллюстрация и текст: BS97 / TEXT / P12 / intro.htm в Медицинской школе Университета Висконсин-Мэдисон
- Совет медицинских исследований
- Береговая лаборатория
Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological:
Behavioural genetics ·
Evolutionary psychology ·
Neuroanatomy ·
Neurochemistry ·
Neuroendocrinology ·
Neuroscience ·
Psychoneuroimmunology ·
Physiological Psychology ·
Psychopharmacology
(Index, Outline)
| Brain: Cochlear nuclei | |
|---|---|
|
|
|
| Dissection of brain-stem. Dorsal view. («Cochlear nucleus» is labeled on left, fifth from the bottom.) | |
|
|
|
| Terminal nuclei of the cochlear nerve, with their upper connections. (Schematic.) The vestibular nerve with its terminal nuclei and their efferent fibers have been suppressed. On the other hand, in order not to obscure the trapezoid body, the efferent fibers of the terminal nuclei on the right side have been resected in a considerable portion of their extent. The trapezoid body, therefore, shows only one-half of its fibers, viz., those which come from the left. 1. Vestibular nerve, divided at its entrance into the medulla oblongata. 2. Cochlear nerve. 3. Accessory nucleus of acoustic nerve. 4. Tuberculum acusticum. 5. Efferent fibers of accessory nucleus. 6. Efferent fibers of tuberculum acusticum, forming the striae medullares, with 6’, their direct bundle going to the superior olivary nucleus of the same side; 6’’, their decussating bundles going to the superior olivary nucleus of the opposite side. 7. Superior olivary nucleus. 8. Trapezoid body. 9. Trapezoid nucleus. 10. Central acoustic tract (lateral lemniscus). 11. Raphé. 12. Cerebrospinal fasciculus. 13. Fourth ventricle. 14. Inferior peduncle. |
|
| Latin | nuclei cochleares |
| Gray’s | subject #187 788 |
| Part of | Medulla |
| Components | |
| Artery | AICA |
| Vein | |
| BrainInfo/UW | hier-717 |
| MeSH | [1] |

The cochlear nuclei are nuclei of the cochlear nerves located in the base of the fourth ventricle of the brain where they act as the first relay station for the [[auditory nerve fibres. They consist of:
- (a) the dorsal cochlear nucleus, corresponding to the tuberculum acusticum on the dorso-lateral surface of the inferior peduncle; and
- (b) the ventral or accessory cochlear nucleus, placed between the two divisions of the nerve, on the ventral aspect of the inferior peduncle.
Anatomy and function
The CN is the first relay station in the auditory system and is located at the dorso-lateral side of the brainstem, spanning the junction of the pons and medulla. Information is brought via the cochlear nerve, also called Cranial nerve VIII, to the CN. The cochlear nucleus can be divided into ventral and dorsal sections (DCN). The ventral part can further be divided into anterior and posterior sections (AVCN and PVCN), which are separated by the incoming auditory nerve fibers.
The auditory nerve fibers form a highly organized system of connections according to their peripheral innervation of the cochlea. Axons from the spiral ganglion cells of the lower frequency innervate the ventral portions of the dorsal cochlear nucleus and the ventrolateral portions of the anteroventral cochlear nucleus. In contrast, the axons from the higher frequency organ of corti hair cells project to the dorsal portion of the anteroventral cochlear nucleus and the uppermost dorsal portions of the dorsal cochlear nucleus. The mid frequency projections end up in between the two extremes, in this way the frequency spectrum is preserved.
At the level of the cochlear nuclei, the input from the two ears, for the most part, remains separated. There is evidence, however, that a significant number of neurons in the CN receives input from the contralateral CN. Just as the inner hair cells are arranged according to the best frequency (BF), so is the cochlea nucleus. This so-called tonotopic organization is preserved because only a few inner hair cells synapse on the dendrites of a nerve cell in the spiral ganglion, and the axon from that nerve cell synapes on only a very few dendrites in the cochlear nuclues.
Each cochlear nucleus has two parts, dorsal (DCN) and ventral (VCN). The Cochlear Nucleus receives input from each spiral ganglion, and also receives input from other parts of the brain. How the inputs from other areas of the brain affect hearing is unknown.
Location and projections from the CN
Projections to the Cochlear Nuclei
The major input to the cochlear nucleus is from the auditory nerve, a part of Cranial nerve VIII (the vestibulocochlear nerve). The auditory nerve fibers form a highly organized system of connections according to their peripheral innervation of the cochlea. Axons from the spiral ganglion cells of the lower frequencies innervate the lateral-ventral portions of the dorsal cochlear nucleus and the ventrolateral portions of the anteroventral cochlear nucleus. In contrast, the axons from the higher frequency organ of corti hair cells project to the dorsal portion of the anteroventral cochlear nucleus and the dorsal-medial portions of the dorsal cochlear nucleus. The mid frequency projections end up in between the two extremes; in this way the frequency spectrum is preserved. In this way, the cochlear nuclei inherit the tone based organization of the cochleae. This so-called tonotopic organization is preserved because only a few inner hair cells synapse on the dendrites of a nerve cell in the spiral ganglion, and the axon from that nerve cell synapes on only a very few dendrites in the cochlear nucleus.
The cochlear nuclei have long been thought to receive input only from the ipsilateral ear. There is evidence, however, for stimulation from the contralateral ear via the contralateral CN,[1] and also the somatosensory parts of the brain.[2]
Projections from the Cochlear Nuclei
There are three major projections from the cochlear nuclei. Through the medulla, one projection goes to the contralateral superior olivary complex (SOC) via the trapezoid body, whilst the other half shoots to the ipsilateral SOC. This projection is called the ventral acoustic stria (or, more commonly, the trapezoid body). Another projection, called the dorsal acoustic stria (DAS, also known as the stria of von Monakow), rises above the medulla into the pons where it hits the nucleus of the lateral lemniscus along with its kin, the intermediate acoustic stria (IAS, also known as the stria of Held). The IAS decussates across the medulla, before joining the ascending fibers in the contralateral lateral lemniscus. The lateral lemniscus contains cells of the nuclei of the lateral lemniscus, and in turn projects to the inferior colliculus. The inferior colliculus receives direct, monosynaptic projections from the superior olivary complex the contralateral dorsal acoustic stria, some classes of stellate neurons of the VCN, as well as from the different nuclei of the lateral lemniscus.
All of these inputs terminate in the inferior colliculus, although there are a few small projections that bypass the inferior colliculus and project to the medial geniculate, or other forebrain structures.
- Medial superior olive (MSO) via Trapezoid Body (TB) – Ipsilateral and contralateral stimulation for low frequency sounds.
- Lateral superior olive (LSO) directly and via TB – Ipsilateral stimulation for high frequency sounds.
- Medial Nucleus of Trapezoid body (MNTB) – Contralateral stimulation.
- Inferior colliculus – Contralateral stimulation.
- Periolivary nuclei (PON) – Ipsilateral and Contralateral stimulation.
- Lateral lemniscus (LL) and Lemniscal Nuclei (LN) – Ipsilateral and Contralateral Stimulation.
Cell types
There are four types of principal cells found in the cochlear nuclei: Bushy cells, stellate cells, octopus cells, and fusiform cells.
- Bushy cells are found in the anterior ventral cochlear nucleus (AVCN). These can be divided into spherical and globular bushy cells, depending on their appearance, and also their location. Within the AVCN here is an area of large spherical cells; caudal to this are smaller spherical cells, and globular cells. They have a few (1-4) very short dendrites with numerous small branching, which cause it to resemble a “bush”. The bushy cells are only found in the ventral portion of the cochlear nucleus. The bushy cells have specialized electrical properties that allow them to transmit timing information from the auditory nerve to more central areas of the auditory system. Some bushy cells can even improve the precision of the timing information. Bushy cells have responses very similar to those in the auditory nerve. The primary difference is that spontaneous activity is decreased by stimulation by adjacent frequencies, therefore leading to an even sharper tuning curve than seen in auditory nerve cells. These cells are usually innervated only by a selected few axons, which dominate its firing patterns. These afferent axons wrap their terminal branches around the entire soma, creating a large synapse onto the bushy cells. Therefore, a single unit recording of an electrically stimulated bushy neuron characteristically produces exactly one action potential and constitutes the primary response.
- Stellate cells, morphologically, have a radial, star-like dendritic tree, which is where they get their name. They are also called, chopper cells, in reference to their ability to fire a regularly-spaced train of action potentials for the duration of a tonal or noise stimulus. The chopping pattern is intrinsic to the electrical excitability of the stellate cell, and the firing rate depends on the strength of the auditory input more than on the frequency.
- Octopus cells are found in a small region of the Posterior Ventral Cochlear Nucleus (PVCN). The distinguishing features of these cells are their long, thick dendrites that typically emanate from one side of the cell body. Octopus cells produce an «Onset Response» to simple tonal stimuli. That is, they respond only at the onset of a specific frequency or frequency range at higher amplitudes. The octopus cells can fire with some of the highest temporal precision of any neuron in the brain. Electrical stimuli to the auditory nerve has been shown to evoke a graded post synaptic potential in the octopus cells. These EPSP’s are very brief. The octopus cells are thought to be important extracting timing information. It has been reported that these cells can respond to click trains at a rate of 800 Hz.
- Fusiform cells (also known as pyramidal cells) are found in the Dorsal Cochlear Nucleus (DCN). See the separate section on the DCN below.
Neurotransmitters: There are four neurotransmitters responsible for transmission of neural impulses, namely, GABA, Glycine, Norepinephrine, Glutamate, and Acetylcholine.
Structures
The input of auditory stimulus is through the Auditory Nerve (CN VIII). The auditory stimulus further travels through the AVCN as neural impulses and branches through the following structures:
- Medial superior olive (MSO) via Trapezoid Body (TB) – Ipsilateral and contralateral stimulation for low frequency sounds.
- Lateral superior olive (LSO) directly and via TB – Ipsilateral stimulation for high frequency sounds.
- Medial Nucleus of Trapezoid body (MNTB) – Contralateral stimulation.
- Inferior colliculus – Contralateral stimulation.
- Neurotransmitters: There are four neurotransmitters responsible for transmission of neural impulses, namely, GABA, Norepinephrine, Glutamate, and Acetylcholine.
- Periolivary nuclei (PON) – Ipsilateral and Contralateral stimulation.
- Lateral lemniscus (LL) and Lemniscal Nuclei (LN) – Ipsilateral and Contralateral Stimulation.
- Inferior colliculus – Contralateral stimulation.
See also
- Auditory system
- Cochlear nerve
References
Young, Eric D., Spirou, George A., Rice, John J., and Voigt, Herbert F., «Neural organization and responses to complex stimuli in the dorsal cochlear nucleus,» Phil. Trans. R. Soc. Lond. B (1992) 336, 407-413
- ↑ Davis KA (September 2005). Contralateral effects and binaural interactions in dorsal cochlear nucleus. J. Assoc. Res. Otolaryngol. 6 (3): 280–96.
- ↑ Shore, S.E. (2009). «Auditory/Somatosensory Interactions» Squire Encyclopedia of Neuroscience, 691–5, Academic Press.
Additional images
")
Dissection of brain-stem. Lateral view.
")
The cranial nerve nuclei schematically represented; dorsal view. Motor nuclei in red; sensory in blue.
")
Nuclei of origin of cranial motor nerves schematically represented; lateral view.
")
Primary terminal nuclei of the afferent (sensory) cranial nerves schematically represented; lateral view.
")
Scheme showing the course of the fibers of the lemniscus; medial lemniscus in blue, lateral in red.
External links
- University of Buffalo
- Neuroanatomy at UW Bs97/TEXT/P12/intro.htm
- Medical research council
This article was originally based on an entry from a public domain edition of Gray’s Anatomy. As such, some of the information contained herein may be outdated. Please edit the article if this is the case, and feel free to remove this notice when it is no longer relevant.
|
v·d·e Human brain, rhombencephalon, metencephalon: pons (TA A14.1.05.101-604, GA 9.785) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dorsal/ (tegmentum) |
|
|||||||||||
| Ventral/ (base) |
|
|||||||||||
| Other grey: Raphe/ reticular |
Reticular formation (Caudal, Oral, Tegmental, Paramedian) · Raphe nuclei (Median) |
|
v·d·e Brain: rhombencephalon (hindbrain) |
||||||||
|---|---|---|---|---|---|---|---|---|
| Metencephalon/ pons |
|
|||||||
| Myelencephalon/ medulla |
|
I-IV: olfactory — optic — oculomotor — trochlear
V: trigeminal: trigeminal ganglion
V1: ophthalmic: lacrimal — frontal (supratrochlear, supraorbital) — nasociliary (long root of ciliary, long ciliary, infratrochlear, posterior ethmoidal, anterior ethmoidal) — ciliary ganglion (short ciliary)
V2: maxillary: middle meningeal — in the pterygopalatine fossa (zygomatic, zygomaticotemporal, zygomaticofacial, sphenopalatine, posterior superior alveolar)
in the infraorbital canal/infraorbital nerve (middle superior alveolar, anterior superior alveolar)
on the face (inferior palpebral, external nasal, superior labial, infraorbital plexus) — pterygopalatine ganglion (deep petrosal, nerve of pterygoid canal)
branches of distribution (palatine, nasopalatine, pharyngeal)
V3: mandibular: nervus spinosus — medial pterygoid — anterior (masseteric, deep temporal, buccal, lateral pterygoid)
posterior (auriculotemporal, lingual, inferior alveolar, mylohyoid, mental) — otic ganglion — submandibular ganglion
VI: abducens
VII: facial: nervus intermedius — geniculate — inside facial canal (greater petrosal, nerve to the stapedius, chorda tympani)
at exit from stylomastoid foramen (posterior auricular, digastric — stylohyoid)
on face (temporal, zygomatic, buccal, mandibular, cervical)
VIII: vestibulocochlear: cochlear (striae medullares, lateral lemniscus) — vestibular
IX: glossopharyngeal: fasciculus solitarius — nucleus ambiguus — ganglia (superior, petrous) — tympanic — carotid sinus
X: vagus: ganglia (jugular, nodose) — Alderman’s nerve — in the neck (pharyngeal branch, superior laryngeal ext and int, recurrent laryngeal)
in the thorax (pulmonary branches, esophageal plexus) — in the abdomen (gastric plexuses, celiac plexus, gastric plexus)
XI: accessory XII: hypoglossal
|
v·d·e Auditory and vestibular pathways |
||||
|---|---|---|---|---|
| Auditory |
inner ear: Hair cells → Spiral ganglion → Cochlear nerve VIII → pons: Cochlear nuclei (Anterior, Dorsal) → Trapezoid body → Superior olivary nuclei → midbrain: Lateral lemniscus → Inferior colliculi → thalamus: Medial geniculate nuclei → cerebrum: Acoustic radiation → Primary auditory cortex |
|||
| Vestibular |
inner ear: Vestibular nerve VIII → pons: Vestibular nuclei (Medial vestibular nucleus, Lateral vestibular nucleus) cerebellum: Flocculonodular lobe spinal cord: Vestibulospinal tract (Medial vestibulospinal tract, Lateral vestibulospinal tract) thalamus: Ventral posterolateral nucleus cerebrum: Vestibular cortex Vestibulo-oculomotor fibers |
|||
| {| class=»navbox collapsible nowraplinks» style=»margin:auto; » | ||||
|
[. v]·[. d]·[. e]
|
||||
|
|}
From Wikipedia, the free encyclopedia
| Cochlear nerve | |
|---|---|

Diagrammatic longitudinal section of the cochlea. (Cochlear nerve is in center, shown as striped.) |
|

Part of the cochlear division of the acoustic nerve, highly magnified. |
|
| Details | |
| From | Vestibulocochlear nerve |
| Identifiers | |
| Latin | nervus cochlearis |
| MeSH | D003056 |
| TA98 | A14.2.01.133 |
| TA2 | 6318 |
| FMA | 53431 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
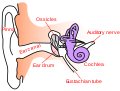
The cochlear nerve (also auditory nerve or acoustic nerve) is one of two parts of the vestibulocochlear nerve, a cranial nerve present in amniotes, the other part being the vestibular nerve. The cochlear nerve carries auditory sensory information from the cochlea of the inner ear directly to the brain. The other portion of the vestibulocochlear nerve is the vestibular nerve, which carries spatial orientation information to the brain from the semicircular canals, also known as semicircular ducts.[1]
Anatomy and connections[edit]
In terms of anatomy, an auditory nerve fiber is either bipolar or unipolar, with its distal projection being called the peripheral process, and its proximal projection being called the axon; these two projections are also known as the «peripheral axon» and the «central axon», respectively. The peripheral process is sometimes referred to as a dendrite, although that term is somewhat inaccurate. Unlike the typical dendrite, the peripheral process generates and conducts action potentials, which then «jump» across the cell body (or soma) and continue to propagate along the central axon. In this respect, auditory nerve fibers are somewhat unusual in that action potentials pass through the soma. Both the peripheral process and the axon are myelinated.
In humans, there are on average 30,000 nerve fibers within the cochlear nerve.[2] The number of fibers varies significantly across species; the domestic cat, for example, has an average of 50,000 fibers. The peripheral axons of auditory nerve fibers form synaptic connections with the hair cells of the cochlea via ribbon synapses using the neurotransmitter glutamate. The central axons form synaptic connections with cells in the cochlear nucleus of the brainstem.
The cell bodies of the cochlear nerve lie within the cochlea and collectively form the spiral ganglion, named for the spiral shape it shares with the cochlea. These central axons exit the cochlea at its base and form a nerve trunk, which, in humans, is approximately one inch long. This travels in parallel with the vestibular nerves through the internal auditory canal, through which it connects to the brainstem. There, its fibers synapse with the cell bodies of the cochlear nucleus.
Types of neurons[edit]
In mammals, cochlear nerve fibers are classified as either type I or type II.
- Type I neurons make up 90-95% of the neurons and innervate the inner hair cells. They have relatively large diameters, are bipolar, and are myelinated. Each type I axon innervates only a single inner hair cell, but each inner hair cell is innervated by up to 30 such nerve fibers, depending on species and location within the cochlea.
- Type II neurons make up the remaining 5-10% of the neurons and innervate the outer hair cells. They have relatively small diameters, are unipolar, and are unmyelinated.
Cochlear nuclear complex[edit]
In mammals, the axons from each cochlear nerve terminate in the cochlear nuclear complex that is ipsilaterally located in the medulla of the brainstem. The cochlear nucleus is the first ‘relay station’ of the central auditory system and receives mainly ipsilateral afferent input.
The three major components of the cochlear nuclear complex are (see figure below):
- the dorsal cochlear nucleus (DCN)
- the anteroventral cochlear nucleus (AVCN)
- the posteroventral cochlear nucleus (PVCN)
Each of the three cochlear nuclei are tonotopically organized. The axons from the low-frequency region of the cochlea project to the ventral portion of the dorsal cochlear nucleus and the ventrolateral portions of the anteroventral cochlear nucleus. The axons from the high-frequency region project to the dorsal portion of the anteroventral cochlear nucleus and the uppermost dorsal portions of the dorsal cochlear nucleus. The axons from the intermediate frequency region project to intermediate targets, such that tonotopy is preserved between the cochlea and the cochlear nuclei.
See also[edit]
- Cochlear implant
- Auditory Brainstem Response
References[edit]
- ^ Palmer, A R (1987). «Physiology of the cochlear nerve and cochlear nucleus». British Medical Bulletin. 43 (4): 838–855. doi:10.1093/oxfordjournals.bmb.a072221. ISSN 1471-8391. PMID 3329928.
- ^ Spoendlin H, Schrott A (1989). «Analysis of the human auditory nerve». Hear Res. 43 (1): 25–38. doi:10.1016/0378-5955(89)90056-7. PMID 2613564. S2CID 37038957.
Additional images[edit]
-

Ear anatomy
-
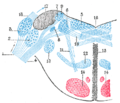
Cochlear nucleus innervated by a branching auditory nerve fibre
-

Terminal nuclei of the vestibular nerve, with their upper connections.
-
Transverse section of the cochlear duct of a fetal cat.
-

Floor of ductus cochlearis.
-

Diagrammatic longitudinal section of the cochlea
-



External links[edit]
- Illustration at cytochemistry.net
- p. 1059
| Кохлеарные ядра | |
|---|---|
|
Рассечение ствола мозга . Вид сзади. («Кохлеарное ядро» указано слева, пятое снизу.) |
|
|
Терминальные ядра улиткового нерва с их верхними связями. (Схема.) Вестибулярный нерв с его конечными ядрами и их эфферентными волокнами подавлен. С другой стороны, чтобы не закрывать трапециевидное тело, эфферентные волокна терминальных ядер с правой стороны были резецированы на значительной части их протяженности. Таким образом, трапециевидное тело показывает только половину волокон, а именно те, которые идут слева.
|
|
| Подробности | |
| Часть | мозговой ствол |
| Система | Слуховая система |
| Артерия | AICA |
| Идентификаторы | |
| латинский | ядра улитки |
| MeSH | D017626 |
| NeuroNames | 720 |
| НейроЛекс ID | birnlex_1151 |
| TA98 |
А14.1.04.247 А14.1.05.430 |
| TA2 | 6006 , 6007 |
| FMA | 72240 |
|
Анатомические термины нейроанатомии
[ редактировать в Викиданных ] |
Кохлеарный ядерные ( CN ) Комплекс состоит из двух черепных нервов ядра в человеческом стволе головного мозга , то вентральные кохлеарные ядра (VCN) и дорсальные кохлеарные ядра (DCN). Вентральное ядро улитки не слоистое, тогда как дорсальное ядро улитки слоистое. Волокна слухового нерва , волокна, которые проходят через слуховой нерв (также известный как кохлеарный нерв или восьмой черепной нерв), переносят информацию от внутреннего уха, улитки , на той же стороне головы, к нервному корню в вентральном ядре улитки. . В корешке нерва волокна разветвляются, иннервируя вентральное ядро улитки и глубокий слой дорсального ядра улитки. Таким образом, вся акустическая информация поступает в мозг через ядра улитки, где начинается обработка акустической информации. Выходы из ядер улитки поступают в более высокие области слухового ствола мозга .
Структура
Ядра улитки (CN) расположены на дорсо-латеральной стороне ствола мозга , охватывая соединение моста и продолговатого мозга .
- Вентральный кохлеарные ядра (VCN) на вентральной части ствола мозга, вентролатерального к нижнему стеблу .
- Дорсальные кохлеарное ядро (DCN), также известный как бугорок acusticum или акустический бугорок, кривые над ОХНОМ и обертывание вокруг мозжечковой ножки.
- VCN далее разделяется нервным корешком на задневентральное ядро кохлеара (PVCN) и антеровентральное ядро кохлеара (AVCN).
Проекции на ядра улитки
Основной вход в кохлеарное ядро поступает от слухового нерва, части VIII черепного нерва ( вестибулокохлеарного нерва ). Волокна слухового нерва образуют высокоорганизованную систему связей в соответствии с их периферической иннервацией улитки. Аксоны спиральных ганглиозных клеток более низких частот иннервируют вентролатеральные части вентрального ядра улитки и латерально-вентральные части дорсального ядра улитки. Аксоны от высокочастотного органа волосковых клеток кортикального слоя проецируются в дорсальную часть вентрального ядра улитки и в дорсально-медиальные части дорсального ядра улитки. Среднечастотные проекции оказываются между двумя крайностями; таким образом тонотопическая организация, установленная в улитке, сохраняется в ядрах улитки. Эта тонотопная организация сохраняется, потому что только несколько внутренних волосковых клеток синапсов на дендритах нервной клетки в спиральном ганглии, а аксон от этой нервной клетки синапсируется только на очень небольшом количестве дендритов в ядре улитки. В отличие от VCN, который получает весь акустический вход от слухового нерва, DCN получает входной сигнал не только от слухового нерва, но также получает акустический вход от нейронов в VCN (Т-звездчатых клетках). Таким образом, DCN в некотором смысле является сенсорным ядром второго порядка.
Долгое время считалось, что ядра улитки получают сигнал только от ипсилатерального уха. Однако есть доказательства стимуляции от контралатерального уха через контралатеральный CN, а также соматосенсорные части мозга.
Проекции ядер улитки
Есть три основных пучка волокон, аксоны нейронов кохлеарного ядра, которые переносят информацию от ядер улитки к мишеням, которые находятся в основном на противоположной стороне мозга. Через продолговатый мозг одна проекция идет к контралатеральному верхнему оливному комплексу (SOC) через трапециевидное тело , а другая половина — к ипсилатеральному SOC. Этот путь называется вентральной акустической полосой (ВАШ или, чаще, трапециевидным телом). Другой путь, называемый дорсальной акустической полосой (DAS, также известной как полоса фон Монакова), поднимается над мозговым веществом в мост, где он поражает ядра латерального лемниска вместе с его родственниками, промежуточной акустической полосой (IAS, также известная как полоса Хельда). IAS пересекает продолговатый мозг перед тем, как присоединиться к восходящим волокнам контралатерального бокового лемниска. Боковой лемниск содержит клетки ядер бокового лемниска и, в свою очередь, выступает в нижний бугорок . Нижний бугорок получает прямые моносинаптические проекции от верхнего оливкового комплекса, контралатеральной дорсальной акустической полоски, некоторых классов звездчатых нейронов VCN, а также от различных ядер латерального лемниска.
Большинство этих входов оканчиваются в нижнем бугорке, хотя есть несколько небольших выступов, которые обходят нижний бугорок и проецируются на медиальный коленчатый вал или другие структуры переднего мозга.
- Медиальная верхняя олива (MSO) через трапециевидное тело (TB) — Ипсилатеральная и контралатеральная стимуляция для звуков низкой частоты.
- Боковая верхняя олива (LSO) напрямую и через TB — ипсилатеральная стимуляция для звуков высокой частоты.
- Медиальное ядро трапециевидного тела (MNTB) — Контралатеральная стимуляция.
- Низший бугорок — контралатеральной стимуляции.
- Периоливные ядра (ПОН) — Ипсилатеральная и контралатеральная стимуляция.
- Боковой лемниск (LL) и лемнисковые ядра (LN) — Ипсилатеральная и контралатеральная стимуляция.
Гистология
Три типа основных клеток передают информацию из вентрального ядра улитки: кустистые клетки, звездчатые клетки и клетки осьминога.
- Кустовидные клетки обнаруживаются в основном в переднем вентральном ядре улитки (AVCN). В дальнейшем их можно разделить на большие сферические, маленькие сферические и шаровидные кустистые клетки, в зависимости от их внешнего вида, а также их расположения. Внутри AVCN есть область больших сферических клеток; каудальнее к нему расположены более мелкие сферические клетки, а глобулярные клетки занимают область вокруг нервного корешка. Важное различие между этими подтипами состоит в том, что они проецируются на разные цели в высшем оливковом комплексе. Крупные сферические густые клетки выступают на ипсилатеральную и контралатеральную медиальную верхнюю оливу. Глобулярные кустистые клетки проецируются на контралатеральное медиальное ядро трапециевидного тела, а маленькие сферические кустистые клетки, вероятно, проецируются на латеральную верхнюю оливу. У них есть несколько (1-4) очень коротких дендритов с многочисленными мелкими ветвлениями, которые делают его похожим на «куст». Кустообразные клетки обладают особыми электрическими свойствами, которые позволяют им передавать информацию о времени от слухового нерва к более центральным областям слуховой системы. Поскольку кустистые клетки получают входной сигнал от нескольких волокон слухового нерва, которые настроены на одинаковые частоты, пушистые клетки могут улучшить точность информации о времени, по сути, усредняя дрожание во времени входных сигналов. Густые клетки также могут подавляться звуками, близкими к частоте, на которую они настроены, что приводит к еще более резкой настройке, чем это наблюдается в волокнах слухового нерва. Эти клетки обычно иннервируются только несколькими волокнами слухового нерва, которые доминируют в его возбуждении. Эти афферентные нервные волокна обвивают свои концевые ветви вокруг всей сомы, создавая большой синапс на густых клетках, называемый « концевой луковицей Хельда ». Следовательно, единичная запись электрически стимулированного кустистого нейрона, как правило, дает ровно один потенциал действия и составляет первичный ответ.
- Звездчатые клетки (также известные как мультиполярные клетки) имеют более длинные дендриты, расположенные параллельно пучкам волокон слухового нерва. Их также называют чопперами, в связи с их способностью запускать последовательность потенциалов действия с регулярным интервалом в течение всего тонального или шумового стимула. Схема прерывания является неотъемлемой частью электрической возбудимости звездчатой клетки, и частота импульсов зависит от силы слухового сигнала больше, чем от частоты. Каждая звездчатая ячейка имеет узкую настройку и имеет тормозящие боковые полосы, что позволяет населению звездчатых клеток кодировать спектр звуков, усиливая спектральные пики и спады. Эти нейроны обеспечивают акустический ввод в DCN.
- Клетки осьминога находятся в небольшой области заднего вентрального ядра улитки (PVCN). Отличительными чертами этих клеток являются их длинные, толстые дендриты в форме щупалец, которые обычно исходят из одной стороны тела клетки. Клетки осьминога вырабатывают «начальную реакцию» на простые тональные стимулы. То есть они реагируют только на появление широкополосного стимула. Клетки осьминога могут активироваться с высочайшей временной точностью среди нейронов мозга. Электрические стимулы слухового нерва вызывают в клетках осьминога ступенчатый возбуждающий постсинаптический потенциал. Эти EPSP очень краткие. Считается, что клетки осьминога важны для извлечения информации о времени. Сообщалось, что эти клетки могут реагировать на последовательность щелчков с частотой 800 Гц.
Два типа основных клеток передают информацию от дорсального кохлеарного ядра (DCN) к контралатеральному нижнему бугорку. Основные ячейки получают две системы входов. Акустический сигнал поступает в глубокий слой несколькими путями. Возбуждающий акустический сигнал поступает от волокон слухового нерва, а также от звездчатых клеток VCN. Акустический вход также передается через тормозящие интернейроны (туберкуловентральные клетки DCN и «широкополосные ингибиторы» в VCN). Через самый внешний молекулярный слой DCN получает другие типы сенсорной информации, в первую очередь информацию о расположении головы и ушей, через параллельные волокна. Эта информация распространяется через мозжечковую цепь, которая также включает тормозящие интернейроны.
- Веретенообразные клетки (также известные как пирамидные клетки ). Веретенообразные клетки интегрируют информацию через два пучка дендритов, апикальные дендриты получают мультисенсорную, возбуждающую и тормозную информацию через самый внешний молекулярный слой, а базальные дендриты получают возбуждающий и тормозящий акустический вход от базальных дендритов, которые распространяются в глубокий слой. Считается, что эти нейроны позволяют млекопитающим анализировать спектральные сигналы, которые позволяют нам локализовать звуки на высоте и когда мы теряем слух на одно ухо.
- Гигантские клетки также интегрируют входящие данные из молекулярного и глубокого слоев, но преобладают входные данные из глубокого слоя. Неясно, какова их роль в слушании.
Функция
Ядерный комплекс улитки — это первая интегративная, или обрабатывающая стадия слуховой системы . Информация доставляется к ядрам из ипсилатеральной улитки через кохлеарный нерв . В ядрах улитки выполняется несколько задач. Распределяя акустический вход на несколько типов основных клеток, слуховой путь подразделяется на параллельные восходящие пути, которые могут одновременно извлекать различные типы информации. Клетки вентрального ядра улитки извлекают информацию, которая переносится слуховым нервом во время возбуждения и в структуре активации популяции волокон слухового нерва. Клетки дорсального кохлеарного ядра выполняют нелинейный спектральный анализ и помещают этот спектральный анализ в контекст расположения головы, ушей и плеч и отделяют ожидаемые, самогенерируемые спектральные сигналы от более интересных, неожиданных спектральных сигналов, используя вход от слуховой коры , ядер моста , ганглия и ядра тройничного нерва, ядер дорсальной колонны и второго ганглия дорсального корешка . Вполне вероятно, что эти нейроны помогают млекопитающим использовать спектральные сигналы для ориентации на эти звуки. Информация используется более высокими отделами ствола мозга для достижения дальнейших вычислительных целей (таких как определение местоположения источника звука или улучшение отношения сигнал / шум ). Поступающие из этих других областей мозга, вероятно, играют роль в локализации звука.
Чтобы понять более подробно специфические функции ядер улитки, сначала необходимо понять, каким образом звуковая информация представлена волокнами слухового нерва . Вкратце, в каждом из двух слуховых нервов около 30 000 волокон слухового нерва. Каждое волокно является аксоном спиральной ганглиозной клетки, которая представляет определенную частоту звука и определенный диапазон громкости. Информация в каждом нервном волокне представлена скоростью потенциалов действия, а также конкретным временем появления индивидуальных потенциалов действия. Конкретная физиология и морфология каждого типа клеток ядра улитки усиливают различные аспекты звуковой информации.
Смотрите также
- Слуховая система
- Кохлеарный нерв
- Униполярная щеточная ячейка
Дополнительные изображения
-
Рассечение ствола мозга. Боковой вид.
-
Схематично изображены ядра черепных нервов; вид со спины. Моторные ядра выделены красным цветом; сенсорный синий.
-
Схематично изображены первичные терминальные ядра афферентных (сенсорных) черепных нервов; вид сбоку.
-
Схема, показывающая ход волокон лемниска; медиальный лемниск синий, латеральный красный.
-
Поперечный разрез нижнего моста, показывающий ядро улитки (# 1), помеченное вверху справа.
Рекомендации
Эта статья включает текст, находящийся в общественном достоянии, со страницы 788 20-го издания «Анатомии Грея» (1918 г.)
Young ED, Spirou GA, Rice JJ, Voigt HF (июнь 1992 г.). «Нейронная организация и ответы на сложные стимулы в дорсальном ядре улитки». Филос. Пер. R. Soc. Лондон. B Biol. Sci . 336 (1278): 407–13. DOI : 10.1098 / rstb.1992.0076 . PMID 1354382 .
внешняя ссылка
- Университет Буффало в веб-архивах Библиотеки Конгресса (архив 27 ноября 2001 г.)
- Иллюстрация и текст: Bs97 / TEXT / P12 / intro.htm в Медицинской школе Университета Висконсин-Мэдисон.
- Совет медицинских исследований
- Береговая лаборатория
| Улитковый нерв | |
|---|---|
| Схематический продольный разрез улитки. (Улитковый нерв находится в центре, показан полосатым.) |
|
| Часть улиткового отдела слухового нерва, сильно увеличена. |
|
| Подробности | |
| Из | Вестибулокохлеарный нерв |
| Идентификаторы | |
| Латинский | nervus cochlearis |
| MeSH | D003056 |
| TA98 | A14.2.01.133 |
| TA2 | 6318 |
| FMA | 53431 |
| Анатомический термины нейроанатомии [редактировать в Викиданных ] |
кохлеарный нерв (также слуховой или слуховой нейрон ) является одной из двух частей вестибулокохлеарный нерв, черепной нерв, присутствующий в амниотах, другая часть — вестибулярный нерв. Улитковый нерв переносит слуховую сенсорную информацию от улитки внутреннего уха непосредственно в мозг. Другая часть вестибулокохлеарного нерва — это вестибулярный нерв, который несет информацию о пространственной ориентации в мозг от полукружных каналов, также известных как полукружные протоки.
Содержание
- 1 Анатомия и связи
- 2 Типы нейронов
- 3 Кохлеарный ядерный комплекс
- 4 См. Также
- 5 Ссылки
- 6 Дополнительные изображения
- 7 Внешние ссылки
Анатомия и связи
С точки зрения анатомии слуховое нервное волокно может быть либо биполярным, либо униполярным, а его дистальная проекция называется, а его центральная проекция называется аксоном ; эти два выступа также известны как «периферический аксон» и «центральный аксон» соответственно. Периферийный отросток иногда называют дендритом , хотя этот термин несколько неточен. В отличие от типичного дендрита, периферический отросток генерирует и проводит потенциалы действия, которые затем «прыгают» по телу клетки (или соме) и продолжают распространяться вдоль центрального аксона. В этом отношении волокна слухового нерва несколько необычны, поскольку потенциалы действия проходят через сому. И периферический отросток, и аксон миелинизированы.
У человека в улитковом нерве в среднем 30 000 нервных волокон. Количество волокон значительно различается у разных видов; у домашней кошки, например, в среднем 50 000 волокон. Периферические аксоны волокон слухового нерва образуют синаптические связи с волосковыми клетками улитки через ленточные синапсы с использованием нейротрансмиттера глутамата. Центральные аксоны образуют синаптические связи с клетками кохлеарного ядра ствола мозга.
Тела клеток улиткового нерва лежат внутри улитки и вместе образуют спиральный ганглий, названный в честь спиральной формы, которую он разделяет с улиткой. Эти центральные аксоны выходят из улитки в ее основании и образуют у человека длину примерно в один дюйм. Он проходит параллельно с вестибулярными нервами через внутренний слуховой проход, через который он соединяется со стволом мозга. Там его волокна синапсируются с телами клеток ядра улитки.
Типы нейронов
У млекопитающих волокна улиткового нерва классифицируются как тип I или тип II.
- Нейроны типа I составляют 90-95% нейронов и иннервируют внутренние волосковые клетки. Они имеют относительно большой диаметр, биполярны и миелинизированы. Каждый аксон типа I иннервирует только одну внутреннюю волосковую клетку, но каждая внутренняя волосковая клетка иннервируется до 30 таких нервных волокон, в зависимости от вида и местоположения в улитке.
- Нейроны типа II составляют оставшиеся 5. -10% нейронов и иннервируют внешние волосковые клетки. Они имеют относительно небольшой диаметр, униполярны и немиелинизированы.
Ядерный комплекс улитки
У млекопитающих аксоны от каждого улиткового нерва оканчиваются в ядерном комплексе улитки, который ипсилатерально расположен в продолговатом мозге ствола мозга.. Кохлеарное ядро является первой «ретрансляционной станцией» центральной слуховой системы и получает в основном ипсилатеральный афферентный сигнал.
Три основных компонента ядерного комплекса улитки (см. Рисунок ниже):
- дорсальное ядро улитки (DCN)
- антеровентральное ядро кохлеара (AVCN)
- задневентральное ядро улитки (PVCN)
Каждое из трех ядер улитки организовано тонотопически. Аксоны из низкочастотной области улитки проецируются в вентральную часть дорсального ядра улитки и вентролатеральные части антеровентрального ядра улитки. Аксоны из высокочастотной области проецируются в дорсальную часть антеровентрального ядра улитки и самые верхние дорсальные части дорсального ядра улитки. Аксоны из области промежуточной частоты проецируются на промежуточные мишени, так что тонотопия сохраняется между улиткой и ядрами улитки.
См. Также
- Кохлеарный имплант
Ссылки
Дополнительные изображения
-
Анатомия уха
-
Кохлеарное ядро, иннервируемое ветвящимся волокном слухового нерва
-
Терминальные ядра вестибулярного отдела нерв, с их верхними связями.
-
Поперечный разрез протока улитки плода кошки.
-
Дно ductus cochlearis.
-
Схематический продольный разрез улитки
-
Кортиев
Внешние ссылки
- Иллюстрация на cytochemistry.net
- Gray’s page # 1059
В связи с
возникновением речи как средства
межличностного общения, слух у человека
играет особую роль, поскольку именно
его слуховая система предназначена
преимущественно для того, чтобы слышать
речь других людей. Потерю слуха нельзя
недооценить. Известно, что глухота или
даже лишь недостаток слуха представляют
собой наиболее серьезную угрозу для
умственного развития ребенка, так как
именно словесное
общение является важнейшим средством
обучения.
Слух требуется и для умения говорить
самому, поэтому глухие от рождения дети
не научаются говорить, так как лишены
слуховых раздражителей. По этой причине
глухота, возникающая до приобретения
речи, относится к особенно серьезным
проблемам, и тугоухие от рождения дети
должны начинать пользоваться слуховым
аппаратом до 1,5 лет.
►Морфология
и физиология периферического отдела
слухового анализатора. Периферическая
часть слухового анализатора анатомически
тесно связана с периферической частью
вестибулярного анализатора – они
располагаются рядом в каменистой части
(пирамиде) височной кости (внутреннее
ухо). Кроме того, они имеют и филогенетическое
«родство» — происходят из одних и тех
же структур в процессе эволюции.
Строение
органа слуха
(рис. 2.1).
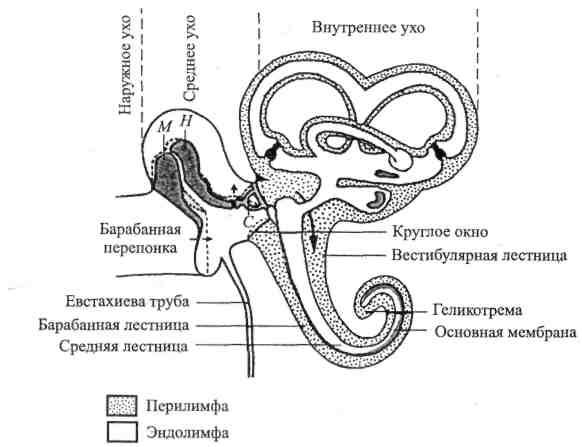
По своему
строению ухо делится на 3 части: 1) наружное
ухо, состоящее из ушной раковины и
наружного слухового прохода; 2) среднее
ухо, состоящее из барабанной полости,
которая находится внутри височной
кости, включает 3 слуховые косточки; 3)
внутренне ухо – самая важная часть
органа слуха.
Рис. 2.1. Схема
среднего и внутреннего уха (по Р. Шмидту
и Г. Тевсу, 1985).
М
– молоточек; Н – наковальня; С –
стремечко. Пунктирной линией показаны
крайние положения косточек под влиянием
колебаний барабанной перепонки.
Звуковые волны
направляются в слуховуюсистему
через наружное
ухо —
наружный слуховой
проход — к барабанной
перепонке. Эта тонкая, с перламутровым
блеском, мембрана замыкает слуховой
проход, образуя
преграду между ним и средним
ухом, в котором также
находится воздух.В
полости среднего уха расположена цепочка
подвижно сочлененных косточек:
молоточек,наковальня истремечко,
что делает более эффективной передачу
звуковых колебаний из воздушного
пространства наружного
слухового прохода в жидкую среду
внутреннего уха. Энергия звука передается
во внутреннее
ухо от барабанной перепонки через
молоточек, наковальню и стремечко,
колеблющиеся
синхронно с ней. «Ручка»
молоточка прочно связана с барабанной
перепонкой, а основание стремечка
(которое действительно выглядит как
стремя) помещено в отверстие каменистой
части височной кости —овальное
окно (здесь
стремечко граничит с внутренним
ухом).
Размеры
площади овального окна
значительно (в 22 раза) меньше воспринимающей
поверхности барабанной перепонки, что
способствует увеличению силы колебаний
барабанной перепонки. Такое различие
в размерах эффективной поверхности
мембраны овального окна и барабанной
перепонки, а также специальная система
рычагов, образованная слуховыми
косточками, создают условия для роста
давления, прилагаемого к овальному
окну, которое примерно в 20-22 раза больше
давления, действующего на барабанную
перепонку. В результате этого даже
слабые звуковые волны, действующие на
барабанную перепонку, способны преодолеть
сопротивление мембраны овального окна
преддверия и привести к колебаниям
жидкости в улитке.
Полость среднего
уха соединяется с глоткой посредством
евстахиевой трубы, которая при
глотании открывается, вентилируя среднее
ухо и уравнивая в нем давление с
атмосферным. Это особенно важно при
резком перепаде давления — при взлете
или посадке самолета, в метро во время
начала движения поезда, в скоростном
лифте или при близкой артиллерийской
стрельбе, когда возникает состояние
«закладывания
ушей». В таком случае необходимостараться путем повышения давления в
полости рта(открыть
рот) или глотания уравнять его с
возрастающим наружным давлением. В
противном случае если это не удается,
возникает опасность разрыва барабанной
перепонки.
Внутреннее ухо
помещается
в каменистой части
височной кости вместе с органом равновесия
и представленокостным лабиринтом,
внутри которого залегаетперепончатый
лабиринт, повторяющий форму костного.
Между обоими лабиринтами имеется
щелевидное пространство, заполненное
перилимфой.Лабиринт состоит из
преддверия, трех полукружных каналов
и улитки.Слуховым органом является
улитка, названная так из-за своей
формы(у человека
она образует 2,5 завитка, у кошки и собаки–
3, у лошади – 2).
Улитка состоит из трех параллельных
свернутых вместе каналов —барабанной
лестницы,
вестибулярной лестницы и
средней лестницы.
Первые два соединяются
между собой у вершины улитки через
маленькое отверстие — геликотрему.
Они заполнены перилимфой,
сходной по химическому составу
с внеклеточной жидкостью и содержащей
поэтому много ионов натрия.
Средняя лестница
заполнена
эндолимфой
– жидкостью,
богатой ионами калия и, стало быть,
сходна с внутриклеточной средой. Пери-
и эндолимфатические области улитки
соединяются с соответствующими областями
вестибулярного
аппарата. Основание стремечка в
овальном окне примыкает к перилимфатическому
пространству вестибулярной лестницы;
отверстие замыкаетсякольцевой
связкой,
так что
перилимфа не может вытекать. В
основании барабанной лестницы имеется
еще одно отверстие, открывающееся
в среднее ухо, — круглое
окно. Оно
также закрыто тонкой мембраной,
предотвращающей вытекание перилимфы.
На
рис. 2.2 показано поперечное сечение
улитки.
Вестибулярная лестница отделяется от
средней лестницы рейснеровой
мембраной,
а основная
(базилярная) мембрана
разделяет
среднюю и
барабанную лестницы. Утолщение,
проходящее вдоль основной мембраны, —кортиев орган,
который
содержит
фонорецепторы,
окруженныеподдерживающими (опорными) клетками.
Выделяют два вида опорных и рецепторных
клеток – наружные и внутренние.
Опорные клетки сходятся под острым
углом друг к другу и образуют заполненный
эндолимфой канал треугольной формы –
внутренний туннель, в котором
расположены нервные волокна, идущие от
нейроновспирального ганглия. Они
также представлены волосковыми клетками,
которые имеют, однако, только стереоцилии;
в рецепторах кортиева органа киноцилии
редуцированы.
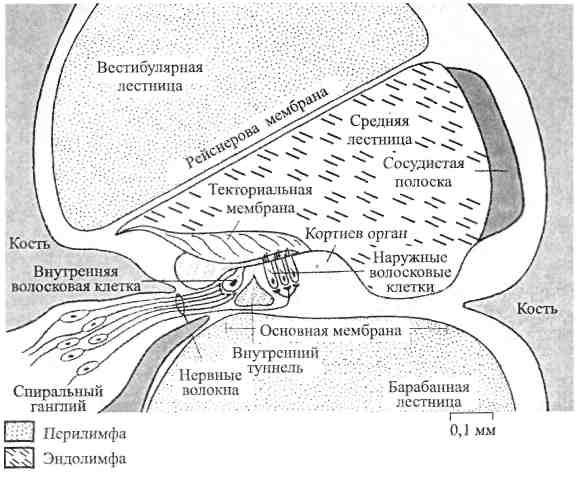
Рис. 2.2. Поперечный
разрез одного из витков спирали улитки
(по Р. Шмидту и Г. Тевсу, 1985).
Фонорецепторы
лежат на опорных клетках. Как и в
вестибулярном аппарате, рецепторные
клетки здесь являются вторичными
сенсорными клетками. На основании их
отношения к кортиеву органу различают
внутренние и наружные волосковые клетки;
последние расположены в три ряда, тогда
как внутренние образуют один. У человекаприблизительно
3500 внутренних и 12000 -20000 наружных
волосковых клеток. На свободной
поверхности фонорецепторов есть волоски,
или стереоцилии (до 100 в клетке). Сегодня
известно, что высокие звуки раздражают
волосковые клетки, расположенные на
нижних завитках улитки, а низкие звуки
возбуждают волосковые клетки вершины
улитки. Показано также, что внутренние
волосковые клетки (в силу их более
высокой чувствительности) лучше отражают
интенсивность звука, чем наружные
волосковые клетки.
Над кортиевым органом лежит текториальная
(покровная) мембрана
(имеет
желеобразную
консистенцию), один край которой
прикрепляется к внутренней стенке
улитки в области центральной кости, а
другой свободно оканчивается в полости
улиткового протока. Стереоцилии
волосковых клеток связаны с нижней
поверхностью текториальной мембраны.
Все фонорецепторы синаптически
связаны с афферентными нервными
волокнами,приходящим
от биполярных клеток спирального
ганглия, который расположенв
центре улитки; центральные отростки
этих клеток
направляются в ЦНС. Около 90% нервных
волокон спирального ганглия оканчиваются
на внутренних волосковых клетках, каждая
из которых образует контакты с множеством
нервных волокон. Только оставшиеся 10%
волокон иннервируют значительно более
многочисленные наружные волосковые
клетки. Чтобы охватить все наружные
клетки, эти волокна должны сильно
разветвляться, хотя иннервируемые
одним волокном клетки могут быть
расположеныблизко
друг к другу. Всего слуховой нерв содержит
примерно от 30 до 40 тысяч афферентных
волокон. Кортиев орган получает
и эфферентные волокна, тормозящие
активность в афферентных волокнах.
На внешней стороне средней лестницы
расположена
сосудистая
полоска
с высокой метаболической активностью
и хорошим кровоснабжением, чтои
отражается в ее названии. Она играет
важную роль вобеспечении улитки
энергией и в регуляции состава
эндолимфы. Различныеионные
насосы, включая калиевый, поддерживают
здесь постоянство ионной средыи
положительный потенциал эндолимфы.
Известно, что некоторые диуретические
препараты являются ототоксичными и
иногда вызывают глухоту. Это обусловлено
тем, что указанные вещества воздействуют
на почки, блокируя ионные насосы в
эпителии канальцев, которые ответственны
за реабсорбцию солей. Очевидно, некоторые
из ионных насосов сосудистой полоски
действуют по сходному принципу и, быть
может, поэтому страдают при применении
этих препаратов.
Механизмы
слуховой рецепции.
Звуковая волна,
воздействуя на систему слуховых косточек
среднего уха, приводит в колебательное
движение мембрану овального окна
преддверия, которая, прогибаясь, вызывает
волнообразные перемещения перилимфы
верхнего (вестибулярной лестницы) и
нижнего (барабанной лестницы) каналов,
которые постепенно затухают по направлению
к вершине улитки. Поскольку все жидкости
несжимаемы, колебания эти были бы
невозможны, если бы не мембрана круглого
окна, которая выпячивается при надавливании
основания стремечка на овальное окно
и принимает исходное положение при
прекращении давления. Колебания перилимфы
передаются на вестибулярную мембрану,
а также на полость среднего канала
(среднюю лестницу), приводя в движение
эндолимфу и базилярную мембрану
(вестибулярная мембрана очень тонкая,
поэтому жидкость в верхнем и среднем
каналах колеблется так, как будто оба
канала едины). При действии на ухо звуков
низкой частоты (до 1000 Гц) происходит
смещение базилярной мембраны на всем
ее протяжении от основания до верхушки
улитки. При увеличении частоты звукового
сигнала происходит перемещение
укороченного по
длине колеблющегося столба жидкости
ближе к овальному окну, к наиболее
жесткому и упругому участку базилярной
мембраны. Деформируясь, базилярная
мембрана смещает стереоцилии волосковых
клеток относительно текториальной
мембраны (они наклоняются). Такое
натяжение чисто механически открывает
от 1 до 5 калиевых каналов в мембране
стереоцилии, в результате чего возрастает
поток ионов калия в стериоцилию (сила
натяжения нити, необходимая для открывания
одного канала, составляет всего около
2 • 10-13
ньютонов). Таким образом, в ответ на
воздействие звука волосковая клетка
деполяризуется,
при этом максимальная величина
рецепторного
потенциала наблюдается уже спустя
100-150 мс от начала воздействия звука.
Однако открывание всего нескольких
ионных каналов в мембране одной
стереоцилии недостаточно для возникновения
рецепторного потенциала достаточной
величины. Для усиления сенсорного
сигнала на рецепторном уровне слуховой
системы предусмотрен специальный
механизм, который заключается в
механическом взаимодействии всех
стереоцилии каждой волосковой клетки.
Дело в том, что все стереоцилии одного
рецептора связаны между собой в пучок
тонкими поперечными нитями. Поэтому,
когда сгибается один или несколько
более длинных волосков, они тянут за
собой все остальные волоски. В результате
этого открываются ионные каналы всех
волосков, обеспечивая достаточную
величину рецепторного потенциала. В
свою очередь этот потенциал, т.е.
деполяризация пресинаптического
окончания волосковой клетки, приводит
к выходу в синаптическую щель
нейромедиатора. Воздействуя на
постсинаптическую мембрану афферентного
волокна,
медиатор вызывает генерацию в нем
возбуждающего
постсинаптического потенциала и далее
(при достижении
критического уровня деполяризации) —
генерацию распространяющихся в нервные
центры импульсов.
►Проводящие
пути слухового анализатора. Вся
информация о звуке, воспринимаемая
улиткой, передается в мозг волокнами
преддверно-улиткового нерва (VIII
пара черепных нервов). Существует
пять основных уровней переключения
восходящих слуховых
волокон: кохлеарные ядра продолговатого
мозга, верхнеоливарный комплекс ядер
продолговатого мозга, нижние бугры
крыши среднего мозга, медиальное
коленчатое тело таламуса и слуховая
зона коры больших полушарий (височные
извилины). Кроме
того, по ходу слухового
пути расположено большое количество
небольших
ядер, в которых осуществляется частичное
переключение восходящих слуховых
волокон.
Выше уже отмечалось,
что первыми нейронами слухового пути
являются биполярные нейроны спирального
ганглия, центральные отростки которых
образуют слуховую, или кохлеарную, часть
нерва — ветвь VIII
пары черепно-мозговых нервов. По этому
нерву информация от волосковых (главным
образом, от внутренних) клеток поступает
к нейронам продолговатого мозга, входящих
в состав кохлеарного (улиткового)
комплекса, т.е. к нейронам второго
порядка. В этот комплекс, лежащий в
области вестибулярного поля ромбовидной
ямки, входят два ядра — дорсальное и
вентральное (которое состоит из двух
отделов — переднего и заднего). Аксон
биполярного нейрона спирального ганглия,
подходя к кохлеарным ядрам, раздваивается
— одна его ветвь идет к дорсальному
ядру, другая — к вентральному. Не
исключено, что волокна, идущие от
апикальной части улитки (т.е. несущие
информацию о низких звуках) преимущественно
достигают
нейронов вентрального ядра, а волокна,
идущие от основания улитки (возбуждаются
высокими звуками) передают
свою импульсацию, главным образом,
нейронам
дорсального ядра кохлеарного комплекса.
Таким образом, для кохлеарных ядер
характерно тонотопическое распределение
информации.
Оба кохлеарных
ядра дают восходящие тракты — дорсальный
и вентральный. Аксоны нейронов дорсального
кохлеарного
ядра, не заходя к нейронам верхней оливы,
сразу же направляются через мозговые
полоски к латеральному лемниску, где
часть из них переключается на нейроны
лемниска (III
нейроны), а часть проходит транзитом
до нейронов нижних бугров четверохолмия
или до нейронов медиального коленчатого
тела.
Аксоны вентрального
кохлеарного ядра сразу же направляются
к варолиеву мосту через трапециевидное
тело к верхней оливе, где находится
верхнеоливарный комплекс (часть волокон
идет к ипсилатеральному комплексу,
часть — к контрлатеральному). В его
состав входят два ядра: латеральное и
медиальное. Медиальное ядро получает
информацию одновременно как от
ипсилатрального, т.е. со своей стороны,
так и от контрлатерального, т.е.
противоположной стороны, кохлеарного
ядра, что обеспечивает формирование
бинаурального слуха уже на уровне
верхней оливы.
Аксоны верхнеоливарнных
нейронов направляются к латеральному
лемниску, где часть из них переключаются
на нейроны этого лемниска (IV
нейроны), а часть проходит транзитом к
нейронам нижних бугров четверохолмия
и к нейронам медального коленчатого
тела, которое является
последним переключательным звеном
восходящего слухового
пути.
Таким образом, от
дорсальных и вентральных кохлеарных
ядер информация в конечном итоге
поступает к нижним буграм четверохолмия
и медальному коленчатому телу. Благодаря
этому звуковая информация используется
для реализации ориентировочного рефлекса
на внезапное звуковое раздражение
(поворот головы в сторону источника
звука), а также для регуляции тонуса
скелетной мускулатуры и формирования
взгляда. Одновременно, от нейронов
медиального коленчатого тела информация
достигает нейронов верхней части
височной доли мозга (поля
41 и 42 по Бродману),
т.е. высших акустических центров, где
совершается корковый анализ звуковой
информации.
Следует
подчеркнуть, что для верхнеоливарного
комплекса,
нижних бугров четверохолмия, медиального
коленчатого
тела, а также для первичных проекционных
зон слуховой коры, т.е. для всех важнейших
слуховых центров
характерна тонотопическая организация
структур. Это отражает
существование принципа пространственного
анализа звуков, позволяющего осуществлять
тонкое частотное различение
на всех этажах слуховой системы.
Чрезвычайно важным
свойством слуховой системы является
билатеральная иннервация структур на
каждом уровне. Впервые она появляется
на уровне верхней оливы и дублируется
на каждом последующем уровне. Это
позволяет реализовать способность
человека и животных оценивать место
расположения источника звука.
Наряду с восходящими
путями в слуховой системе имеются и
нисходящие пути, обеспечивающие контроль
высших
акустических центров над получением и
обработкой
информации в периферическом и проводниковом
отделах слухового анализатора. Нисходящие
пути слухового анализатора начинаются
от
клеток слуховой коры, переключаются
последовательно в
медиальных коленчатых телах, нижних
буграх четверохолмия, верхнеоливарном
комплексе, от которого идет оливокохлеарный
пучок Расмуссена, достигающий волосковых
клеток улитки. Кроме того, имеются
эфферентные волокна, идущие от первичной
слуховой зоны, т.е. от височной
области, к структурам экстрапирамидной
двигательной системы
(базальным ганглиям, ограде, верхним
буграм четверохолмия,
красному ядру, черной субстанции,
некоторым
ядрам таламуса, ядрам основания моста,
ретикулярной формации ствола мозга) и
пирамидной системы. Эти данные
указывают на участие слуховой сенсорной
системы в регуляции двигательной
активности человека.
►Переработка
информации в коре больших полушарий.
В повседневной жизни мы почти никогда
не сталкиваемся с чистыми тонами. Звуки,
которые мы слышим, построены из разных
частотных компонентов, которые постоянно
и независимо меняются (человек воспринимает
звуковые колебания с частотой 16-20000 Гц).
Амплитуда таких звуков также может
меняться, как и их частота, варьирует и
их длительность, они также могут
начинаться и заканчиваться резко и
постепенно, их источник может располагаться
ближе, дальше или перемещаться и т.д.
Конечно, в полном объеме люди способны
оценивать все эти свойства звука, только
если их слух достаточно тренирован. Так
вот, нейронные процессы, лежащие в основе
такой оценки, выявляются в основном в
слуховой
области коры (средняя часть верхней
височной извилины).
Так, например, одни нейроны первичной
слуховой коры отвечают только на начало
звукового стимула, а другие – только
на его окончание. Некоторые другие
возбуждаются только при звуках
определенной длительности либо при
повторяющихся звуках. Есть нейроны,
возбуждение которых возникает только
в случае, если стимул определенным
образом модулирован по частоте или
амплитуде. Многие нейроны активируются
при действии звуков в широкой полосе
частот, т.е. шумов, а другие, так называемые
простые нейроны, — вычленяют информацию
о чистых звуках. Есть нейроны, которые
возбуждаются только на определенную
последовательность звуков или на
определенную амплитудную их модуляцию.
Есть нейроны, которые позволяют определить
направление звука. Повреждение височных
долей мозга, т.е. именно тех, где
располагается слуховая кора, затрудняет
восприятие речи, пространственную
локализацию источника звука и идентификацию
временных характеристик звука. Подобные
повреждения, однако, не влияют на
способность различать частоты и
интенсивность звука (см. выше).
Большинство
корковых нейронов активируется
контралатеральным ухом, некоторые
отвечают на ипсилатеральную стимуляцию,
а остальные – только при одновременной
стимуляции того и другого уха. Значительный
процент нейронов первичной слуховой
коры не возбуждаются ни при каких
экспериментальных воздействиях.
Предполагается, что эти нейроны являются
высокоспецифичными и отвечают только
на стимулы, которые сложно воспроизвести
в лабораторных условиях. В целом ответы
клеток первичной слуховой коры сходны
с ответами групп или ансамблей нейронов
зрительной коры.
В целом, в первичных
и вторичных проекционных зонах слуховой
коры происходит сложнейший анализ
звукового сигнала. Однако важной является
и функция ассоциативных зон коры больших
полушарий. Например, представление о
мелодии возникает именно благодаря
деятельности этих зон коры, в том числе
на основе информации, хранящейся в
памяти. Именно с участием ассоциативных
зон коры (с помощью специализированных
нейронов типа «бабушкиных» нейронов)
человек способен максимально извлечь
информацию, поступающую
от различных рецепторов, в том числе и
от фонорецепторов.
Кохлеарный нерв
15.12.2020

Кохлеарный нерв (также слуховой или акустический нейрон) — это одна из двух частей вестибулокохлеарного нерва. Улитковый нерв переносит слуховую сенсорную информацию от улитки внутреннего уха непосредственно в мозг. Другая часть вестибулокохлеарного нерва — это вестибулярный нерв, который несет информацию о пространственной ориентации в мозг из полукружных каналов, также известных как полукружные протоки. Данный нерв проходит совместно с вестибулярным нервом в составе VIII пары черспномозговых нервов через внутренний слуховой проход и мостомозжечковое пространство до входа в ствол мозга. Волокна кохлеарного нерва в продолговатом мозгу направляются к своим кохлеарным ядрам (дорсальному и вентральному).
Анатомия
С точки зрения анатомии, волокно слухового нерва бывает биполярным или униполярным, причем его дистальная проекция называется периферическим отростком, а центральная проекция — аксоном. Эти два выступа также известны как «периферический аксон» и «центральный аксон» соответственно. Периферический отросток иногда называют дендритом, хотя этот термин несколько неточен. В отличие от типичного дендрита, периферический отросток генерирует и проводит импульсы, которые затем перемещаются по телу клетки (или соме) и продолжают распространяться вдоль центрального аксона.
У людей в кохлеарном нерве в среднем 30 000 нервных волокон. Количество волокон значительно различается у разных видов: у домашней кошки, например, в среднем 50 000 волокон. Периферические аксоны волокон слухового нерва образуют синаптические связи с волосковыми клетками улитки через ленточные синапсы. Центральные аксоны образуют синаптические связи с клетками кохлеарного ядра ствола мозга. Тела клеток улиткового нерва лежат внутри улитки и вместе образуют спиральный ганглий, названный так из-за спиральной формы, которую он разделяет с улиткой. Эти центральные аксоны выходят из улитки в ее основании и образуют нервный ствол, который у человека составляет приблизительно один дюйм в длину. Он проходит параллельно с вестибулярными нервами через внутренний слуховой проход, через который он соединяется со стволом мозга. Там его волокна синапсируют с телами клеток ядра улитки.
Типы нейронов
У млекопитающих кохлеарные нервные волокна классифицируются как тип I или тип II.
Нейроны типа I составляют 90-95% нейронов и иннервируют внутренние волосковые клетки. Они имеют относительно большой диаметр, биполярны и миелинизированы. Каждый аксон типа I иннервирует только одну внутреннюю волосковую клетку, но каждая внутренняя волосковая клетка иннервируется до 30 таких нервных волокон, в зависимости от вида и местоположения внутри улитки. Нейроны типа II составляют оставшиеся 5-10% нейронов и иннервируют внешние волосковые клетки. Они имеют относительно небольшой диаметр, униполярны и немиелинизированы.
Кохлеарный ядерный комплекс
У млекопитающих аксоны каждого улиткового нерва оканчиваются ядерным комплексом улитки, который ипсилатерально расположен в продолговатом мозге ствола. Кохлеарное ядро является первой «ретрансляционной станцией» центральной слуховой системы и получает в основном ипсилатеральный афферентный сигнал. Каждое из трех ядер улитки тонотопически организовано. Аксоны из низкочастотной области улитки проецируются в вентральную часть дорсального ядра улитки и вентролатеральные части антеровентрального ядра улитки. Аксоны из высокочастотной области проецируются в дорсальную часть антеровентрального ядра улитки и самые верхние дорсальные части дорсального ядра улитки. Аксоны из области промежуточной частоты проецируются на промежуточные мишени, так что тонотопия сохраняется между улиткой и ядрами улитки.
Смотрите также
Кохлеарный имплант
- Только сильнейшие
- Паульсен, Луи
- Древлехранилище ЯГУАК
- Турессон, Гёте Вильгельм
- Коффинит
- Испанская синагога
- Кехтна (волость, 2017)
- Упрощённая система налогообложения
- Амедей I (граф Невшателя)
- Теорема сравнения Штурма