Мозжечок.Структурно-функциональная организация.
Мозжечок
представляет собой образование,
расположенное позади больших полушарий
мозга над продолговатым мозгом и мостом
мозга. Это одна из интегративных структур
головного мозга, принимающая участие
в координации и регуляции произвольных,
непроизвольных движений, в регуляции
вегетативных и поведенческих функций.
Реализация указанных
функций обеспечивается следующими
морфологическими особенностями мозжечка:
-
кора
мозжечка построена достаточно однотипно,
имеет стереотипные связи, что создает
условия для быстрой обработки информации; -
наиболее
крупным и функционально-значимым
элементом среди нейронов коры мозжечка
является клетка Пуркинье. Она имеет
большое количество входов и формирует
единственный аксонный выход из мозжечка,
коллатерали которого заканчиваются
на его ядерных структурах; -
на
клетки Пуркинье проецируются практически
все виды сенсорных раздражений:
проприоцептивные, кожные, зрительные,
слуховые, вестибулярные и др.; -
выходы
из мозжечка обеспечивают его связи с
корой большого мозга, стволовыми
образованиями и спинным мозгом.
Мозжечок анатомически
и функционально делится на старую,
древнюю и новую части.
К
старой части мозжечка (вестибулярный
мозжечок) – относится клочково-флоккулярная
доля. Эта часть имеет наиболее выраженные
связи с вестибулярным анализатором,
что объясняет значение мозжечка в
регуляции равновесия.
Древняя
часть мозжечка (спинальный
мозжечок) – состоит из участков червя
и пирамиды мозжечка, язычка, околоклочкового
отдела и получает информацию преимущественно
от проприоцептивных систем мышц,
сухожилий, надкостницы, оболочек
суставов. Используя полученные сигналы
спинальный мозжечок принимает участие
в регуляции мышечного тонуса и контроле
движений преимущественно мышц конечностей
и аксиальных мышц тела.
Новый
мозжечок включает
в себя кору полушарий мозжечка и участки
червя.Он получает информацию от коры,
преимущественно по лобно-мостомозжечковому
пути, от зрительных и слуховых рецептирующих
систем, что свидетельствует об его
участии в анализе зрительных, слуховых
сигналов и организации на них реакции.
Кора мозжечка
имеет специфическое, нигде в ЦНС не
повторяющееся строение.
Верхний
(1) слой
коры мозжечка – молекулярный
слой,
состоит из параллельных волокон,
разветвлений дендритов и аксонов 2 и 3
слоёв. В нижней части молекулярного
слоя встречаются корзинчатые и звёздчатые
клетки, которые обеспечивают взаимодействие
клеток Пуркинье.
Средний
(2) слой
коры образован клетками Пуркинье,
выстроенными в один ряд и имеющими самую
мощную в ЦНС дендритную систему. На
дендритном поле одной клетки Пуркинье
может быть до 100 тыс. синапсов. Эти клетки
выполняют сбор, обработку и передачу
информации. Аксоны клеток Пуркинье
являются единственным путем, с помощью
которого кора мозжечка передает
информацию в его ядра и ядра структуры
большого мозга.
Под
2 слоем коры (под клетками Пуркинье)
лежит гранулярный
(3) слой,
состоящий из клеток-зёрен, число которых
достигает 10 млрд. Аксоны этих клеток
поднимаются вверх, Т-образно делятся
на поверхности коры, образуя дорожки
контактов с клетками Пуркинье; здесь
же лежат клетки Гольджи.
В
кору мозжечка от кожных рецепторов,
мышц, суставных оболочек, надкостницы
сигналы поступают по так называемым
спиномозжечковым трактам: заднему
(дорсальному) и переднему (вентральному).
Эти пути к мозжечку проходят через
нижнюю оливу продолговатого мозга. От
клеток олив идут так называемые лазающие
(лиановидные) волокна,
которые ветвятся на дендритах клеток
Пуркинье.
Я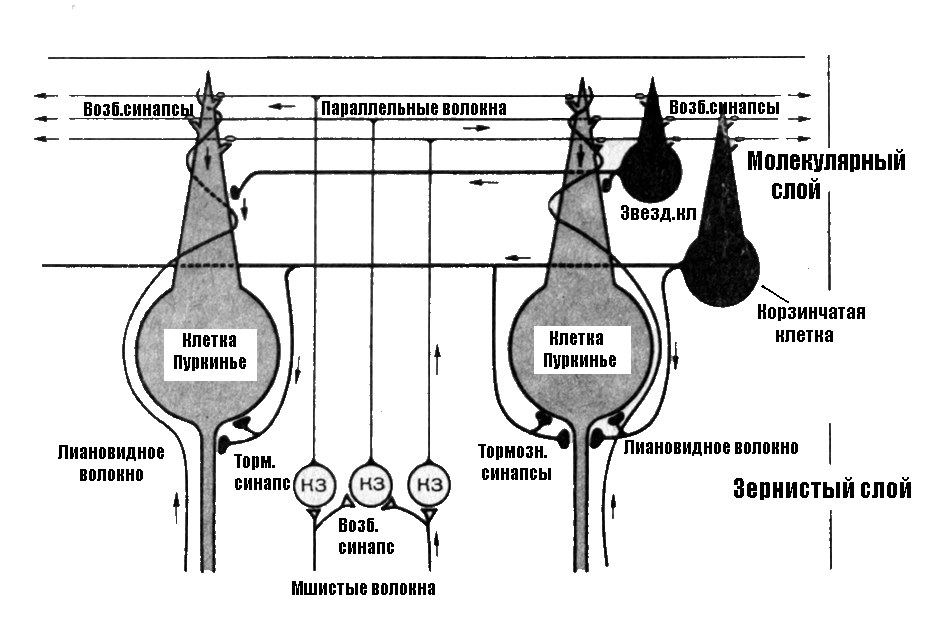 дра
дра
моста посылают афферентные пути в
мозжечок, образующие мшистые
(моховидные) волокна,
которые оканчиваются на клетках-зернах
III
слоя коры мозжечка.
Между
мозжечком и голубым ядром среднего
мозга существует афферентная связь с
помощью адренергических волокон. Эти
волокна способны диффузно выбрасывать
норадреналин в межклеточное пространство
коры мозжечка и за счет этого изменять
возбудимость его клеток.
Лиановидные
волокна непосредственно взаимодействуя
с клетками Пуркинье (медиатор аспартат),
оказывают на них возбуждающее влияние,
но действуя через корзинчатые (медиатор
ГАМК) и звёздчатые (медиатор таурин)
клетки поверхностного слоя коры мозжечка
оказывают на них (более слабое) тормозное
действие.
Моховидные
волокна образуют возбуждающие синапсы
на дендритах клеток-зёрен внутреннего
слоя коры. Через аксоны клеток-зёрен
моховидные волокна оказывают возбуждающее
влияние (медиатор глутамат) на клетки
Пуркинье и опосредованно тормозное
действие через корзинчатые и звёздчатые
клетки. Активность клеток-зёрен
регулируется через тормозные клетки
Гольджи внутреннего слоя по типу
возвратного торможения (медиатор ГАМК).
Таким
образом, на клетки Пуркинье поступает
преимущественно возбуждающее афферентное
влияние. Но, поскольку клетки Пуркинье
являются тормозными нейронами (медиатор
ГАМК), их возбуждение оказывает тормозное
влияние на подкорковые структуры
мозжечка. Следовательно, мозжечок
осуществляет регуляцию не по механизму
запуска нейронной активности моторных
центров, а по механизму более или менее
сдерживания этой активности.
Подкорковая
система мозжечка состоит из четырёх
функционально разных ядерных образований:
ядра шатра,
пробковидного, шаровидного и зубчатого
ядер.
Ядро
шатра получает информацию от медиальной
зоны коры мозжечка и связано с ядром
Дейтерса и ретикулярной формацией
продолговатого и среднего мозга. Отсюда
сигналы идут по ретикулоспинальному
пути к мотонейронам спинного мозга.
Промежуточная
кора мозжечка проецируется на пробковидное
и шаровидное ядра. От них связи идут в
средний мозг к красному ядру, далее в
спинной мозг по руброспинальному пути.
Второй путь от промежуточной коры
мозжечка идет к таламусу и далее в
двигательную зону КБП.
Зубчатое
ядро, получая информацию от латеральной
зоны коры мозжечка, связано с таламусом,
а через него с моторной зоной КБП.
Из
мозжечка информация уходит через верхние
и нижние ножки. Через верхние ножки
сигналы идут в таламус, в ретикулярную
формацию среднего мозга, ядра ствола
мозга. Через нижние ножки мозжечка
сигналы идут в продолговатый мозг к
оливам, ретикулярной формации. Средние
ножки мозжечка связывают новый мозжечок
с лобной долей мозга.
Функции мозжечка.
Несмотря
на исключительную упорядоченность
нейронной организации мозжечка его
функциональная
роль раскрыта еще не полностью.
Клинические
проявления, развивающиеся при поражении
мозжечка, а также эффекты,
наблюдаемые при его раздражении или
экстирпации, свидетельствуют о важной
роли мозжечка
в осуществлении статических,
статокинетических рефлексов и
других процессов
управления двигательной активностью,
автоматически регулирующих работу
двигательного
аппарата.
Экспериментальные
и клинические наблюдения показывают,
что при поражениях мозжечка развиваются
разнообразные нарушения двигательной
активности и мышечного тонуса,
а также вегетативные расстройства.
Основным проявлением расстройств
двигательной
системы является классическая триада
симптомов (триада
Лючиани):астения,
атаксия и атония.
Астения
– быстрая утомляемость и снижение силы
мышц.
Атаксия
– нарушение координации движений. У
таких больных неуверенная, шаткая
походка с широко расставленными ногами,
отведёнными балансирующими руками, с
помощью которых больной поддерживает
равновесие тела. Движения неуверенные,
сопровождающиеся избыточными
толчкообразными бросками из стороны в
сторону. Больной не может стоять и ходить
на носках или пятках.
Атония–
снижение тонуса мышц и недостаточность
поддержания позы тела. После
исследования
коленного рефлекса или смещения пассивно
висящей конечности нога не возвращается
в исходное положение, а раскачивается
подобно маятнику.
Полное
удаление мозжечка или его передней доли
у животных приводит к повышению
тонуса мышц-разгибателей, в то время
как раздражение передней доли–
к снижению этого тонуса.
Также
характерными проявлениями мозжечковой
недостаточности являются:
—тремор
–небольшие
по амплитуде колебательные движения,
возникающие синхронно в различных
участках тела. Мозжечковый тремор
отмечается не в состоянии покоя, а во
время движения. При целенаправленных
действиях амплитуда периодических
движений становиться столь размашистой,
что мешает достигать цели движения
(пальценосовая проба);
—
астазия
–
невозможность сохранять спокойное
состояние;
—
асинергия
–нарушение
взаимодействия между двигательными
центрами
различных мышц. Так, у больных при
выполнении движений не происходит
одновременного
сокращения мышц, компенсирующих смещение
центра тяжести. Делая шаг,
больной выносит ногу, не сгибая, впереди
туловища, что приводит к падению. В
результате
асинергии сложные движения как бы
распадаются на ряд выполняемых
последовательно
более простых движений.
—
адиадохокинез–
замедление скорости смены одной
двигательной реакции на другую (сгибание
и разгибание);
—
дистония
– непроизвольное повышение или понижение
тонуса мышц. Локальные
повреждения небольших участков коры
передней доли мозжечка позволили выявить
локализацию в ней представительства
различных участков скелетной мускулатуры;
—
дизартрия–
дефект речи, при котором речь больного
становиться замедленной, невнятной,
нечленораздельной (наблюдается при
двустороннем повреждении коры мозжечка);
—
скандированная
речь–
дефект речи, при котором больной говорит
медленно, произнося излишне раздельно
не только слова, но и слоги.
Нарушение
двигательной координации при поражениях
мозжечка объясняется тесными связями
его с основными структурами ствола
мозга (дающими начало трактам, передающим
импульсы в спинной мозг), а также с
таламусом и сенсомоторной областью
коры больших полушарий.
Таким
образом, мозжечок активно участвует в
поддержании тонуса мышц и позы, поддержании
равновесия тела в пространстве,
программировании предстоящих движений
и их осуществлении, участие в организации
координации сложных движений. Мозжечок
играет важную роль в процессах моторного
обучения.
Средний
мозг. Структурно-функциональная
организация.
Средний
мозг, так же, как и продолговатый мозг
и мост мозга (варолиев мост), относятся
к стволовым
структурам. Он
занимает относительно небольшую по
объему часть мозгового ствола. Входящие
в его состав образования в ходе эволюции
подвергались значительно большим
изменениям, чем структуры заднего мозга.
Анатомически
средний мозг состоит из двух основных
компонентов:
дорсального
отдела,
обозначаемого как крышка
мозга,
и
вентрального — ножки
мозга.
В
составе
среднего мозга выделяют:
-
ядра
черепных нервов (III и IV
пары); -
бугры четверохолмия;
-
красное ядро;
-
черную субстанцию;
-
голубое ядро;
-
ретикулярную
формацию; -
через
средний мозг проходят различные
восходящие
пути к
таламусу мозжечку, нисходящие
пути.
Функции
среднего мозга. Рефлекторная
деятельность среднего мозга.
Средний
мозг принимает участие в выполнении
сложных и разнообразных рефлекторных
функций, осуществление которых происходит
при участии его структур.
К одной из структур
среднего мозга относятся черепно-мозговые
нервы, ядра которых локализованы в
среднем мозге:
IV
пара
– блоковый
нерв.Являетсядвигательным.Иннервирует
верхнюю косую мышцу, которая поворачивает
глазное яблоко кнаружи и вниз.
III
пара –глазодвигательный
нерв.Являетсядвигательным.
Аксоны
мотонейронов глазодвигательного
нерва иннервируют верхнюю, нижнюю и
внутреннюю косую мышцы глаза, а
также мышцу, поднимающую веко. Нейроны
добавочного ядра глазодвигательного
нерва (ядро Якубовича, относится к
парасимпатическому отделу ВНС) регулируют
просвет зрачка и кривизну хрусталика;
непарное мелкоклеточное ядро Перлиа
иннервирует цилиарную мышцу и участвует
в актах аккомодации и конвергенции.
Рефлекторные
функции осуществляются функционально
самостоятельными структурами среднего
мозга – буграми
четверохолмия.
В нижнем двухолмии располагаются
нейроны, получающие сигналы по слуховым
путям
(первичный
слуховой центр). В
верхнем двухолмии располагаются клетки,
на которые
переключается импульсация, приходящая
по зрительным путям (первичный
зрительный
центр). От
нейронов бугров четверохолмия аксоны
идут к ретикулярной формации ствола,
мотонейронам спинного мозга.
Основная
функция бугров четверохолмия
– организация реакции настораживания
и так называемых «старт-рефлексов» на
внезапные, еще не распознанные, зрительные
или звуковые сигналы.Активация среднего
мозга в этих случаях через гипоталамус
приводит к повышению тонуса мышц,
учащению сердцебиения; происходит
подготовка к оборонительной реакции.В
случае повышенной возбудимости
четверохолмий при внезапном звуковом
или световом раздражителе человек
вздрагивает, иногда вскакивает на ноги,
вскрикивает и максимально быстро
удаляется от раздражителя. При повреждении
четверохолмий эти реакции затормаживаются.
Таким
образом, ядра четверохолмия принимаю
участие вформировании ориентировочных
рефлексови организации произвольных
движений.
Из
клеточных групп среднего мозга особенно
выделяются красное ядро и черная
субстанция.
Красное
ядро
содержит нервные клетки разных размеров.
От крупных нейронов начинаются
наиболее толстые и быстропроводящие
аксоны руброспинального тракта. Эти
нейроны получают сигналы возбуждающего
характера из моторной зоны коры, из
промежуточного ядра мозжечка, а также
из нервных клеток черной субстанции.
Нейроны
красного ядра участвуют в регуляции
распределения тонуса скелетных мышц и
движений, обеспечивающих сохранение
нормального положения тела в пространстве
и принятие позы, создающей готовность
к выполнению определённых действий.
Основа влияний – руброспинальный тракт,
волокна которого оказывают возбуждающее
влияние на α-
и γ-мотонейроны
мышц – сгибателей и тормозят большинство
мотонейронов мышц – разгибателей.
Красное
ядро организовано соматотопически:
клетки, аксоны которых иннервируют
спинальные
центры, контролирующие мускулатуру
верхних и нижних конечностей, начинаются
из различных зон красного ядра.
Децеребрационпая
ригидность.
После перерезки ствола мозга
(децеребрации)ниже
уровня красного ядра
у животных наблюдается выраженное
изменение тонуса мускулатуры туловища
и конечностей — децеребрационная
ригидность, т.е.
резкое
повышение тонуса мышц-разгибателей.
Конечности при этом сильно вытянуты,
голова запрокинута назад, хвост приподнят.
Развитие
децеребрационной ригидности связано
с выключением импульсации, поступающей
в спинной мозг по кортикоспинальному
и руброспинальному трактам, которые
активируют в первую очередь мотонейроны
мышц-сгибателей. При этом начинает
преобладать активность вестибулоспинальной
системы, повышающей тонус преимущественно
мотонейронов мышц-разгибателей.
У
человека ригидность может наблюдаться
при повреждении вышерасположенных
структур ствола мозга и подкорковых
ядер без повреждения среднего мозга.
При этом у человека она проявляется
усилением
тонуса мышц-сгибателей.

Важным
образованием среднего мозга является
черная
субстанция
— скопление нервных
клеток, содержащих пигмент меланин,
придающий этому ядру характерный темный
цвет. Ее нейроны получают многочисленные
проекции от нервных клеток базальных
ганглиев.
В свою очередь они образуют синаптические
связи с нейронами ретикулярных ядер
ствола мозга, ядрами таламуса и базальными
ганглиями.
Черная
субстанция участвует в сложной координации
движений, регуляции тонуса мышц и позы
тела. В ней сосредоточены
содержащие дофамин
нейроны,
многие из которых посылают аксоны в
передний мозг. Они
принимают участие в регуляции
эмоционального поведения. Другая часть
дофаминергических
нейронов черной субстанции посылает
аксоны к ядрам полосатого тела (оказывая
преимущественно тормозное влияние),
где дофамин играет важную роль в контроле
сложных двигательных актов. Повреждения
черной субстанции, приводящие к
дегенерации дофаминергических волокон,
проецирующихся в полосатое тело, приводят
к нарушению тонких движений пальцев
рук, развитию
мышечной ригидности и тремору (болезнь
Паркинсона).
Голубое
ядро
представляет собой плотное скопление
нейронов, отростки которых образуют
дивергентные сети с одним входом. Нейроны
пятна обеспечивают норадренергическую
иннервацию большинства областей ЦНС.
Оно имеет связи с корой больших полушарий,
таламусом и гипоталамусом, мозжечком,
спинным мозгом, и принимает участие в
обеспечении гомеостаза, регуляции
циклов сна и бодрствования. Голубое
ядро участвует в формировании эмоциональных
реакций. Недостаток норадреналина
в нейронах ядра сопровождается
возникновением депрессии, избыток –
дистрессовыми состояниями.
Проводниковая
функция среднего мозга.
Через
средний мозг проходят все восходящие
пути к таламусу (медиальная петля и
спинноталамический путь), мозжечку и
КБП. Нисходящие пути через средний мозг
идут к продолговатому и спинному мозгу.
Это пирамидный путь, корково-мостовые
волокна, руброретикулоспинальный путь.
Средний
мозг находится в подчинении вышележащих
структур ЦНС и сам управляет нижележащими
отделами.
Ретикулярная
формация ствола мозга. Структурно-функциональная
организация и роль в интегративной
деятельности ЦНС.
Р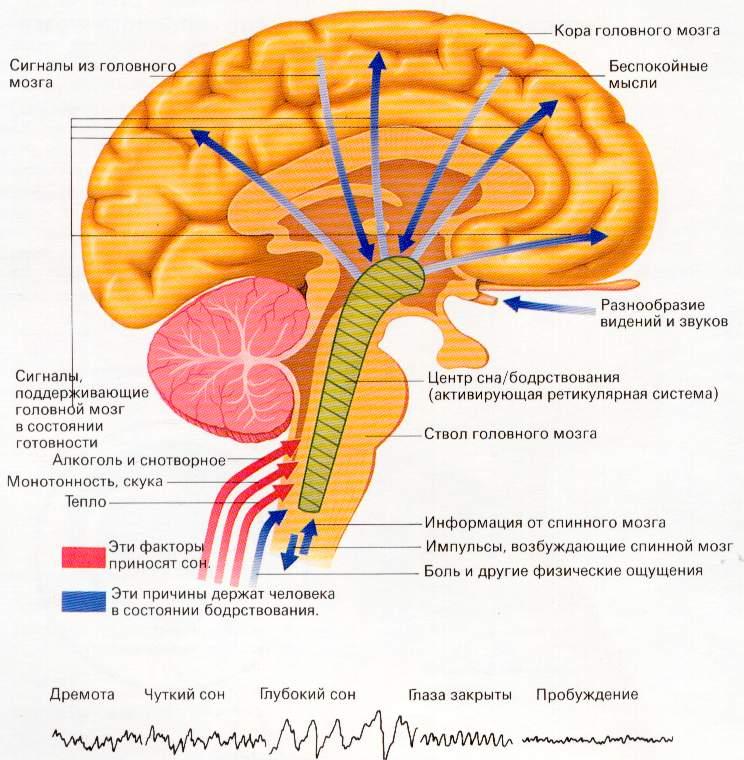 етикулярная
етикулярная
формация (РФ)—
это связанный практически со всеми
структурами ЦНС комплекс полиморфных
нейронов различных размеров с огромным
количеством коллатералей и отростков,
между которыми имеются тесные контакты
в виде химических и электрических
синапсов, расположенных от спинного
мозга до неспецифических ядер таламуса.
Впервые ретикулярная формация была
описана в 1885 году Дейтерсом, который и
дал ей название.
Особенности
нейронов ретикулярной формации (РФ):
-
большинство
нейронов имеет длинные дендриты и
короткий аксон; -
активность
нейронов РФ различна: существуют нейроны
с постояннойритмической активностью
и «молчащие» нейроны; -
на
нейроны РФ конвергируют нервные импульсы
от восходящих и нисходящих специфических
путей, проходящих по стволу мозга и
дающих сюда коллатерали; -
нейроны
РФ высоко чувствительны к химическим
воздействиям (они легко блокируются
снотворными средствами, барбитуратами,
аминозином); -
нейроны
РФ полисенсорны, т.е. возбуждаются на
раздражения, поступающие от различных
рецепторов.
Ретикулярная
формация имеет афферентные
входы преимущественно
от трёх источников: 1) от температурных
и болевых рецепторов по волокнам
спиноретикулярного пути и тройничного
нерва; 2) от сенсорной и частично других
зон коры головного мозга по
кортикоретикулярным путям; 3) от ядер
мозжечка по мозжечковоретикулярному
пути.
Эфферентные
выходы:
1) в спинной мозг – по латеральному и
медиальному ретикулоспинальным путям;
2) к верхним отделам головного мозга
(неспецифическим ядрам таламуса,
гипоталамусу, полосатому телу) от
ретикулярных ядер продолговатого мозга
и моста; 3) к мозжечку от ядер покрышки
моста.
До 1935 года учёные
думали, что хаотически разбросанные
по стволу мозга нейроны в виде сетчатого
образования не выполняют никакой
функции. Но в 1935 году Бремер
поставил опыты с перерезками ствола
мозга у кошки и вживлением микроэлектродов
выше перерезки.
Первая
перерезка
производилась между передними, и задними
буграми четверохолмия, в результате
чегоБремервыключал
основную массу РФ, и кошка засыпала. При
этом на ЭЭГ (электроэнцефалограмме)
записывался α-ритм. При перерезке ниже
среднего мозга (т.е. при сохранении
основной массы РФ) кошка бодрствует и
на ЭЭГ пишется β-ритм. На основании этих
опытов был сделан вывод, что РФ выполняет
очень важную физиологическую
функцию:поддерживает
кору в бодрствующем состоянии.
Если РФ выключена, наступает сон.
Ретикулярная
формация исполняет роль фильтра и
пропускает в вышележащие отделы только
новую и важную информацию.
Активность
ретикулярной формации поддерживается
постоянно на высоком уровне т.к. через
нее проходят импульсы от всех рецепторов.
Согласно современным
представлениям различают
восходящие
и нисходящие
влияния РФ.
Восходящие
влияния
обычно носят активирующий характер
(Бремер, 1935). РФ повышает тонус коры и
регулирует возбудимость её нейронов,
не изменяя специфики ответов на адекватные
раздражители. Иногда может наблюдаться
торможение коры, так как кора через РФ
сама регулирует свою активность.
Нисходящее
влияние
открыто И.М. Сеченовым (1862) в опыте с
кристалликом хлорида натрия, помещённым
на таламус лягушки. Он получил торможение
сгибательного рефлекса, определяемого
по методу Тюрка. Однако только в 40-е годы
стало понятно, что кристаллик хлорида
натрия тормозит сгибательный рефлекс
по
механизму возбуждения ретикулярной
формации ствола
и включению тормозящих влияний на
мотонейроны спинного мозга. В настоящее
время установлено (Г. Мегун, Д. Моруцци),
что
нисходящее
влияние РФ оказывает
модулирующее воздействие
на нейроны спинного мозга, т.е. оно может
быть как тормозным, так и облегчающим.
Нисходящее
тормозное влияние:
возбуждение нейронов РФ в медиальной
части продолговатого
мозгамоносинаптическизатормаживает
α-мотонейроны передних рогов спинного
мозга, а также возбуждает тормозные
клетки Реншоу, которые в свою очередь
будут затормаживать α-мотонейроны.
Нисходящее
облегчающее влияние
начинается с нейронов РФ, расположенных
в промежуточном мозге, среднем мозге,
варолиевом мосту и некоторых отделах
продолговатого мозга (но в основном это
ростральный отдел ствола мозга). Это
влияние облегчает возбуждение
α-мотонейронов, угнетает возбуждение
тормозных нейронов Реншоу, оказывает
регулирующее влияние на γ-мотонейроны,
которые изменяют возбуждение мышечных
веретён через интрафузальные мышечные
волокна. Тем самым РФ регулирует тонус
мышц.
Соседние файлы в папке Методички к занятиям
- #
- #
- #
- #
- #
- #
- #
- #
- #
Обновлено: 04.04.2023
Функция мозжечка состоит, в первую очередь, в формировании двигательных программ, необходимых для поддержания равновесия тела, регуляции силы мышц и объема совершаемых с их участием движений.
Он не является составной частью иерархической сети двигательных систем, но имеет связи со всеми ее уровнями: моторной корой, двигательными центрами ствола и спинным мозгом. Благодаря этим связям мозжечок может сравнивать моторные программы коры с их исполнением, а затем корректировать движения, не соответствующие намеченной программе. Мозжечок способен накапливать опыт двигательной активности, сохраняя его в долговременной моторной памяти. Функциональная организация мозжечка
В мозжечке имеются три области, различающиеся спецификой своих соединений с головным и спинным мозгом и выполняемыми функциями: 1) вестибулоцеребеллум (червь с примыкающими к нему областями старой коры); 2) спиноцеребеллум (примыкающие к червю медиальные области полушарий мозжечка); 3) цереброцеребеллум, включающий в себя латеральные области полушарий мозжечка (рис. 4.25). Вестибулоцеребеллум получает афферентную информацию от вестибулярных ядер и к ним же посылает эфферентные сигналы: его основная задача состоит в сохранении равновесия при стоянии и ходьбе, а также в управлении движениями глаз. Спиноцеребеллум основную часть сенсорной информации получает от проводящих афферентных путей спинного мозга — двух спиномозжечковых трактов, несущих импульсы от проприоцепторов мышц и сухожилий. Эфферентная информация от ядер этой части мозжечка поступает к двигательным центрам ствола, относящимся как к медиальному, так и к латеральному нисходящим путям. Спиноцеребеллум контролирует правильность начинающихся движений ног и рук. Цереброцеребеллум получает при участии ядер моста информацию о планирующемся движении из сенсомотор- ных областей коры и посылает эфферентные сигналы к первичной и вторичной моторной коре, участвуя в планировании движений.
> Сохранение равновесия и движения глаз
alt=»» />
Вестибулоцеребеллум (функция сохранения равновесия и управления движениями глаз) получает зрительную и слуховую информацию, входные сигналы от вестибулярных ядер, ядер тройничного нерва и спинного мозга.
Спиноцеребеллум (управление движениями конечностей) получает афферентную информацию от проприоцепторов посредством спиномозжечковых трактов; эфферентный выход к первичной и вторичной моторной коре.
Цереброцеребеллум (формирование двигательных программ) получает афферентную информацию при посредничестве ядер моста от сенсомоторных областей коры; основной эфферентный выход к первичной и вторичной моторной коре.
Таким образом, мозжечок постоянно получает информацию о планирующихся движениях от коры, о положении головы и глаз, о тонусе мышц, необходимом для совершения движения,— от двигательных центров ствола, а от спинного мозга к нему поступают сведения о характере уже совершаемых движений. Располагая всей полнотой информации о движении — от замысла до исполнения, мозжечок постоянно сравнивает совпадение замысла с исполнением. При несоответствии хода движения намеченному плану мозжечок немедленно исправляет возникающие ошибки. Он может вносить коррективы как в двигательную программу, благодаря своим связям с моторной корой, так и в исполнение движения, действуя на двигательные центры ствола, на медиальные и латеральные нисходящие пути.
При выполнении запрограммированных произвольных движений выходная активность нейронов зубчатых ядер мозжечка регистрируется приблизительно на 10 мс раньше, чем она обнаружится в моторной коре. Это опережение имеет особенное значение при выполнении быстрых движе
При мозжечковом поражении пациент, например, промахивается, когда его просят быстро прикоснуться к кончику своего носа указательным пальцем. Взаимодействие нейронов коры и ядер мозжечка
Афферентная информация, адресованная мозжечку, распределяется между его ядрами и корой. Под влиянием афферентных сигналов нейроны ядер мозжечка создают основную часть его выходной активности, и лишь самая старая часть коры — клочок имеет непосредственную связь с вестибулярными ядрами. Функция коры мозжечка состоит в непрерывной модуляции выходной активности ядер тормозными нейронами Пуркинье, использующими в качестве медиатора ГАМК (рис. 4.26).
Во внутреннем слое трехслойной коры мозжечка расположены зернистые клетки, являющиеся входными нейронами коры, а также тормозные по отношению к ним клетки Гольджи. Афферентные моховидные волокна, образованные нейронами нескольких ядер ствола и спинного мозга, передают информацию зернистым клеткам от проприо- цепторов скелетных мышц, вестибулярного аппарата и коры. Аксоны зернистых клеток направлены в наружный слой коры мозжечка, где они Т-образно делятся на длинные волокна, которые возбуждают нейроны Пуркинье и одновременно тормозные интернейроны, сдерживающие активность клеток Пуркинье. Нигде в ЦНС больше не встречается столь выраженное, как в коре мозжечка, преобладание торможения над возбуждением, поэтому поступающее в кору мозжечка возбуждение прекращается уже через 100 мс: так быстро «стирается» информация об осуществлении актов движения — она не должна мешать поступлению новых оперативных данных.
Грушевидные нейроны Пуркинье формируют средний слой коры, они имеют аксоны, образующие синапсы с нейронами ядер мозжечка. Каждая клетка Пурки-
нье получает сигналы от приблизительно 200 000 параллельных волокон зернистых клеток, а каждая зернистая клетка собирает афферентные входы от десятка моховидных волокон.
Еще один афферентный вход в мозжечок представлен лазающими волокнами, которые образованы нейронами нижнего оливарного ядра продолговатого мозга и доставляют информацию от коры и от спинного мозга. Лазающие волокна обертывают тела и дендриты клеток Пуркинье, образуя с ними многочисленные синапсы, причем каждое лазающее волокно контактирует с несколькими (от одного до десяти) нейронами Пуркинье, но каждая клетка Пуркинье получает возбуждение только от одного лазающего волокна. Синапс между окончанием лазающего волокна и нейроном Пуркинье является одним из наиболее эффективных в ЦНС потому, что в ответ на одиночный потенциал действия, проведенный лазающим волокном, в нем возникает высокоамплитудный ВПСП. Результатом этого становится высокочастотный залп потенциалов действия клетки Пуркинье, направленный на нейроны ядер мозжечка и тормозящий их активность. Этот тормозной выход из коры мозжечка является конечным и единственным результатом ее деятельности.
Нейронная сеть коры мозжечка организована по принципу функциональных модулей, каждый из которых занимает сагиттальную зону шириной 1—2 мм. В каждый модуль входит лазающее волокно из определенной части оливы, а каждой клетке Пуркинье достается обособленная информация от моховидных и лазающего волокон. Лазающие волокна модулируют активность моховидных волокон, они могут усиливать их влияние, но могут и ограничивать эффективность определенных входов. В функциональном отношении это означает, что активность одного пути будет изменяться в зависимости от активности другого пути. Соотношение активности отдельных лазающих и моховидных волокон может меняться вследствие приобретаемого двигательного опыта — в результате подобной модуляции синаптических переключений повышается эффективность совершаемых действий. Эфферентные связи мозжечка с моторными структурами мозга
Клетки ядер мозжечка представляют собой обычные переключательные нейроны: в ответ на поступающую по коллатералям моховидных и лазающих волокон афферентную информацию они возбуждаются и отправляют эфферентные сигналы двигательным ядрам ствола.
Клетки Пуркинье ограничивают их активность таким образом, что выполнение одних моторных программ допускается, а других — отменяется в соответствии с характером афферентной информации и организацией связей между корой мозжечка и ядрами.
Эфферентное влияние зубчатых и промежуточных ядер мозжечка на сенсомоторную кору больших полушарий осуществляется при участии моторных вентролатералъных ядер таламуса, которые служат узловым пунктом нейронной сети, связывающей сенсомоторную кору не только с мозжечком, но и с базальными ганглиями. Аксоны таламических нейронов способны изменять активность пирамидных клеток, образующих кортикоспинальный путь, что позволяет мозжечку корректировать двигательные программы, создаваемые в моторных областях коры.
Еще одним пунктом переключения моторных систем мозга являются красные ядра, служащие источником образования руброспинальных путей. Здесь оканчиваются аксоны возбуждающих нейронов промежуточных ядер мозжечка и моторной коры, оказывающей тормозное влияние. От ядер шатра эфферентные волокна идут к вестибулярным ядрам и моторным ядрам ретикулярной формации ствола мозга, что позволяет мозжечку контролировать нисходящую активность вестибулоспинальных и ретикулоспи- нальных путей. Действие мозжечка на стволовые двигательные центры позволяет исправлять ошибки уже запущенной двигательной программы, если они возникают в процессе ее осуществления.
Функции мозжечка
Мозжечок регулирует силу и точность мышечных сокращений и их тонус как в покое, так и при движениях, а также синергию сокращений разных мышц при сложных движениях. При поражении мозжечка возникает целый ряд как двигательных расстройств, так и нарушений со стороны вегетативной нервной системы. К ним относятся астения —быстрая утомляемость, снижение силы мышечных сокращений,астазия —утрата способности к Длительному сокращению мышц, поэтому больной не может долго стоять или сидеть, неточность движений;атаксия —нарушение координации движений, неуверенная походка;абазия —невозможность сохранить центр тяжести тела;атония или дистония — лонижение или повышение тонуса мышц;тремор —дрожание пальцев рук, кистей и головы в покое, но его усиление при движении;дизартрия —нарушение кординации мышц лица, необходимая для четкого произнесения слов. Речь становится монотонной, медленной и невыразительной.Дисметрия —расстройство равномерности движения (гиперметрия или гипометрия).
Страдает кожная и проприоцептивная чувствительность нижних (пучок Флексига) и верхних конечностей (пучок Говерса)- При поражении мозжечка тормозится процесс обучения, так как многие движения человек заучивает во время жизни (письмо, ходьба). I
Нарушение функций мозжечка приводит к неточности движений, их разбросанности, негармоничности, а это говорит об участии мозжечка в деятельности коры больших полушарий, ответственной за организацию процессов высшей нервной деятельности.
Мозжечок влияет на возбудимость сенсомоторной коры больших полушарий и контролирует тем самым уровень тактильной, температурной и зрительной чувствительности.
Мозжечок играет адаптационно-трофическую роль в регуляции не только мышечной деятельнсти, но и ее вегетативного обеспечения. Удаление мозжечка приводит к нарушению целого ряда вегетативных функций (Л. А. Орбели). Мозжечок оказывает как угнетающее, так и стимулирующее влияние на работу сердечно-сосудистой системы. При раздражении последнего высокое артериальное давление снижается, а исходное низкое —повышается. Снижается частота дыхания, повышается тонус гладких мышц кишечника. При повреждении мозжечка нарушается углеводный, белковый и минеральный обмен, а также процессы энергообразования, терморегуляции и кроветворения. Стимуляция мозжечка приводит к нарушению репродуктивной функции, к сокращению матки у беременных кошек. Удаление мозжечка провоцирует рождение нежизнеспособного потомства или препятствует зачатию.
Промежуточный мозг
В состав промежуточного мозга входят: 1)таламус (область зрительногобугра), 2)гипоталамус(подталамическая область)и3)третий желудочек.
Область зрительного бугра, в свою очередь, слагается: 1)из зрительного бугра (таламус оптикус), 2)надталамической области (эпиталамус —эпифиз, шишковидная железа), 3)заталамической области (метаталамус —медиальное и латеральное коленчатые тела).
Таламус
Зрительный бугор является местом переключения всех чувствительных проводников, идущих от экстеро-, проприо- и интерорецепторов, поднимающихся в кору головного мозга. В нем происходит обработка всей информации, поступающей в кору из спинного мозга и подкорковых структура По мнению А.Уолкера. выдающегося исследователя зрительного бугра, «таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым корковым центрам таким образом, чтобы организм смог адекватно приспособиться к постоянно меняющейся среде. Таламус, как видно, таит в себе тайну многого из того, что происходит в коре больших полушарий».
О полифункциональности таламуса говорит наличие в нем около 120ядер, которые топографически можно разделить на три основные группы:переднюю,имеет проекции в поясную кору, медиальную —в лобную,латеральную —в теменную, височную, затылочную.
По функциональным признакам ядра зрительного бугра делят на три группы: специфические,образующие с соответствующими областями коры специфическую таламокортикальную систему,неспецифические,составляющие диффузную, неспецифическую таламокортикальную систему, иассоциативные.
В состав группы специфических ядервходят переднее вентральное, медиальное, вентролатеральное, постмедиальное, постлатеральное, а также медиальные и латеральные коленчатые тела. Специфические ядра содержат так называемые «релейные» (передаточные) нейроны, имеющие мало дендритов и длинный аксон, заканчивающийся вIII— IVслоях коры больших полушарий (соматосенсорная зона).
Если раздражать какое-либо из специфических ядер электрическими импульсами, то в соответствующих проекционных областях коры с коротким латентным периодом возникает реакция, названная первичным ответом.
Каждое из специфических ядер отвечает за свой вид чувствительности, так как они, так же как и кора больших полушарий, имеют соматотопическую локализацию, т.е. к ним поступают сигналы от тактильных, болевых, температурных, мышечных рецепторов, а также от интерорецепторов зон проекции блуждающего и чревного нервов.
Латеральное, или наружное, коленчатое тело —этоподкорковый центр зрения,таламическое реле для зрительных импульсов. Оно имеет афферентные связи с сетчаткой глаза и буграми четверохолмия и эфферентные —с затылочной долей коры больших полушарий.
Медиальное коленчатое тело —подкорковый, таламический Центр слуха,получает афферентные импульсы из латеральной петли и нижних бугров четверохолмия и посылает информацию в ьисочную долю коры больших полушарий.
К неспецифическим ядрамталамуса относятся: срединный Центр, парацентральное ядро, центральное медиальное и латеральное, субмедиальное, вентральное переднее, парафасцикулярное, ретикулярное ядро, перивентрикулярное и центральная серия масса.
Нейроны неспецифических ядер являются клетками ретикулярной формации, аксоны которых контактируют со всеми слоями коры больших полушарий, образуя диффузные связи. В свою очередь, к неспецифическим ядрам поступает информация от ретикулярной формации ствола мозга, лимбической системы, базальных ганглиев и специфических ядер таламуса.
Раздражение неспецифических ядер электрическим током вызывает возникновение в коре больших полушарий не локально, а диффузно специфической электрической активности, имеющей длинный латентный период и вид веретена, названной сонными веретенами, или реакцией вовлечения.
Ассоциативные ядраталамуса включают в себя медиодорсальное, латеральное дорсальное ядро и подушку. Нейроны этих ядер имеют разную форму и количество отростков, что позволяет им выполнять разнообразные функции, связанные с переработкой информации различных модальностей, после чего она поступает в 1-е и 2-е слои ассоциативной зоны коры, частично —в проекционные зоны коры (4-е и 5-е слои).
При поражении таламических ядер, отвечающих за переработку всей сенсорной информации, в том числе и болевой, могут возникать сильнейшие боли. С наличием застойного очага возбуждения в таламусе и коре больших полушарий связаны «фантомные боли» (в ампутированной конечности).
Таламус обеспечивает двигательные и вегетативные реакции, связанные с сосанием, жеванием, глотанием и смехом.
Функции мозжечка
Удаление мозжечка вызывает такие симптомы, как нарушение чувства равновесия, нарушение тонуса скелетной мускулатуры, кроме того, оно приводит к характерным изменениям в осуществлении произвольных движений.
Первые подробные описания признаков нарушений, которые появляются после удаления мозжечка у животных, были сделаны итальянским физиологом Л. Лючиани в 1893 году. Он установил, что после удаления мозжечка наблюдаются три переходящие друг в друга стадии нарушения движений. На первой стадии происходят изменения, зависящие не только от удаления мозжечка, но и от травм, сопутствующих операции, при которой повреждаются связанные с мозжечком отделы мозга, возникают внутричерепные кровоизлияния и отеки ткани. На второй стадии выпадает ряд функций, в осуществлении которых участвует мозжечок. На третьей стадии происходит некоторая компенсация выявленных нарушений. В первые дни после удаления мозжечка животное не может встать на лапы. Затем движения частично восстанавливаются, но остаются беспорядочными. Животное шатается, падает, совершает много лишних, размашистых и неточных движений. Для устойчивости животное широко расставляет лапы. Если у животного удалена только одна половина мозжечка, то конечности соответствующей стороны тела у него вытянуты и при попытке встать оно заваливается на бок или начинает ходить по кругу (манежные движения).
После того как первые тяжелые явления проходят и наступает частичная компенсация функций, животное начинает вставать, ходить, однако расстройства движений той половины тела, на которой удален мозжечок, у него сохраняются.
Человек, у которого наблюдаются мозжечковые расстройства, не может стоять с закрытыми глазами, движения его не координированы. Из-за нарушения координации мышц-антагонистов он не в состоянии несколько раз подряд согнуть и разогнуть любую из конечностей. То есть удаление или поражение мозжечка вызывает расстройство статических и статокинетических рефлексов, а также нарушение произвольных движений. Это свидетельствует о том, что влияние мозжечка распространяется, с одной стороны, на центры ствола мозга, участвующие в осуществлении установочных рефлексов и тонических рефлексов положения тела, с другой стороны, на моторную зону коры больших полушарий, участвующих в организации произвольных движений. Лючиани описал три симптома, характерных для поражения мозжечка: атонию, астению и астазию.
Атония (от греч. tonos — напряжение) — это резкое понижение тонуса мышц, которое возникает через несколько дней после удаления мозжечка. В первое время после операции тонус некоторых мышц, особенно мышц-разгибателей, бывает резко повышен, но через несколько дней он резко падает. Однако согласно наблюдениям Л. А. Орбели, еще через некоторое время тонус некоторых групп мышц может вновь оказаться повышенным. Поэтому считается, что в результате удаления мозжечка происходит не только атония, но и дистония, то есть нарушение регуляций мышечного тонуса.
Астазия (от греч. stasis — стояние) — неспособность стоять, проявляющаяся в появлении качательных и дрожательных движений. При этом состоянии теряется способность (без контроля зрения) правильно поддерживать вертикальную позу; для сохранения равновесия приходится широко расставлять ноги. Голова, туловище и конечности больного непрерывно дрожат и качаются.
Астения (от греч. astheneia — бессилие) — это легкая утомляемость вследствие повышения обмена веществ, связанного с тем, что движения производятся неэкономично, при участии большого количества мышечных групп.
В дальнейшем были описаны и другие симптомы, возникающие при нарушении функций мозжечка.
Атаксия (от греч. ataxia — беспорядок) заключается в недостаточной координации движений, в нарушении их точности. Походка человека становится неровной, зигзагообразной, ноги заплетаются. Его размашистые и слишком резкие движения напоминают движения пьяного человека.
Дисметрия (от греч. metron — мера, размер) — это избыточность или недостаточность амплитуды целенаправленного движения. Нарушение точности, скорости и направления движений.
Нарушение координации произвольных движений легко проверяется, когда больному предлагают закрыть глаза и быстро прикоснуться кончиком указательного пальца к кончику его собственного носа. Если функции мозжечка нарушены, сделать это чрезвычайно трудно. Проявлением нарушения координации произвольных движений является и потеря способности к быстрой смене фаз движений. Например, при разрушении мозжечка быстрые повороты кисти руки в противоположные стороны становятся практически невозможными.
Нарушение мозжечковых функций при дальнейшей жизни высших животных достаточно быстро и эффективно компенсируются за счет того, что новый мозжечок, который у них особенно хорошо развит, имеет обильные корково-мозжечковые связи. При постепенном возникновении поражения мозжечка (например, при медленном росте опухолей, разрушающих мозжечковую ткань) нарушения в некоторых случаях успевают компенсироваться по мере развития повреждения и в результате вообще не проявляются до самой смерти. Такая компенсация обеспечивается, скорее всего, за счет функции коры больших полушарий. Это говорит о том, что мозжечок можно рассматривать как структуру, в какой-то степени дублирующую активность коры при выполнении ею двигательных функций. То есть если в системе «моторная кора — мозжечок» постепенно выходит из строя один из компонентов, то двигательная функция продолжает осуществляться структурой, не захваченной поражением.
Кроме того, Э. А. Асратяном было показано, что нарушения функций после удаления мозжечка могут компенсироваться и благодаря возникновению новых условно-рефлекторных связей в коре полушарий мозга. Если в период относительной компенсации двигательных функций, произошедшей вследствие обучения, у животных без мозжечка удалить моторную зону коры, у них вновь возникнут столь же резкие нарушения состояния двигательного аппарата, как и в первое время после экстирпации мозжечка.
Физиология и функции мозжечка
Физиология и функции мозжечка
Анатомия мозжечка
Филогенетические и функциональные отделы можно рассматривать вместе (приблизительно), если разделить мозжечок на функциональные области. Клочково-узелковая доля также известна как вестибулярный мозжечок, поскольку поступившая к ней информация направляется в вестибулярные ядра; через вестибулярный аппарат она контролирует движения глаз (также она получает информацию от ствола мозга и через мост — от теменной и затылочной коры, что вносит свой вклад в координацию движений глаз).
Отделы червя (дорсальные) получают сигналы от ретикулярной формации, лобных зрительных полей, верхнего холмика и направляют их к глубоким мозжечковым ядрам (ядрам шатра мозжечка) в составе белого вещества, расположенного около узелка червя мозжечка От ядер шатра мозжечка волокна проходят к центрам взора ствола мозга и вестибулярным ядрам, контролирующим саккадические движения глаз.
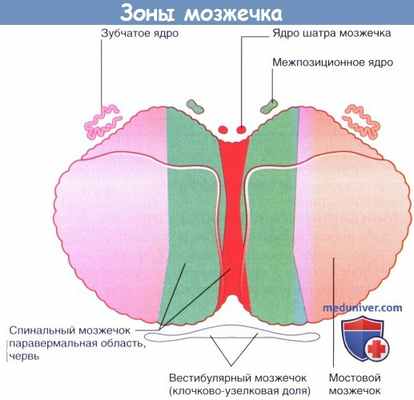
Зоны мозжечка. Внутримозжечковые ядра показаны отдельно.
Парамедианная долька, спинальный мозжечок, включающая червь и паравермальную (прилежащую к червю) кору, получает информацию через спино-мозжечковые пути (кроме того, от вестибулярных ядер и ретикулярной формации, а также через мост — от коры больших полушарий). Червь передает сигналы к ядрам шатра мозжечка, а через волокна — к ретикулярной формации и вестибулярным ядам по ретикулоспинальному и вестибулоспинальному путям контролирует постуральные рефлексы головы и туловища.
К паравермальной области по спиномозжечковым путям подходят волокна от коры больших полушарий через мост и спинной мозг и отходят волокна к шаровидному к пробковидному ядрам. Эти два ядра объединены под названием межпозиционное ядро. Промежуточное ядро взаимодействует с красным ядром и таламусом, которые через волокна, соединяющие их со спинным мозгом (красноядерно-спинномозговой путь) и корой, контролируют и корректируют движения нижних конечностей.
Оставшаяся латеральная долька — самая крупная, посылает информацию через зубчатое ядро. Эту область также обозначают как мостовой мозжечок, поскольку к ней подходит большое количество волокон от противоположных ядер моста. Кроме того, ее называют новой частью мозжечка (неоцеребеллум), так как она получает информацию через ядра моста от обширных областей неокортекса больших полушарий (филогенетически наиболее поздних).
Неоцеребеллум значительно развит в человеческом мозге и играет огромную роль в планировании, начале движения, контроле и коррекции произвольных движений.
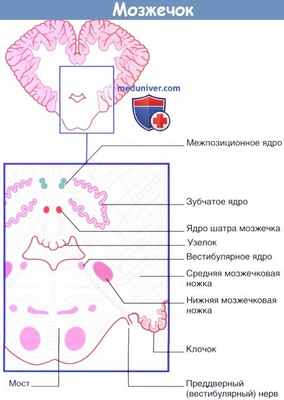
На поперечном срезе через нижний отдел моста и мозжечок показано расположение внутримозжечковых и вестибулярных ядер.
а) Предупреждающая функция мозжечка. Мозжечок выполняет сложные функции стабилизации позы и поддержания позы, описанные ниже.
1. Стабилизация позы. Поражение передней доли приводит к невозможности контролирования по ретикуло-спинномозговым волокнам изменений центра тяжести при ходьбе, что обусловливает нарушения походки.
2. Поддержание позы. На рисунке ниже показан эксперимент, в котором пациента требуют выполнить резкое разгибание в запястье и удержать его в разогнутом состоянии в течение 2 с в момент проведения электромиографии (ЭМГ) первичных разгибателей запястья (короткого и длинного лучевых разгибателей запястья) и главной мышцы-антагониста (лучевого сгибателя запястья). Результаты показали, что мышца-антагонист начала сокращаться до завершения движения, что привело к появлению осцилляций в мыш-цах-синергистах в период фиксации. Действие мышцы-анта-гониста направлено на предотвращение спонтанных колебательных движений (тремора), обусловленных вязкоупругими характеристиками мышц. Было показано, что эти нормальные и необходимые осцилляции могут быть блокированы при транскраниальной электромагнитной стимуляции верхней мозжечковой ножки или при поражении латеральной доли мозжечка.
Упреждающее сокращение икроножной мышцы для стабилизации туловища перед произвольным сокращением двуглавой мышцы плена.
Проще говоря, смещение верхнего отдела туловища от центра тяжести при произвольном движении головы или верхней конечности ожидается мозжечком.
Получив инструкции от премоторных областей лобной коры относительно предполагаемого движения, мозжечок осуществляет пропорциональные сокращения постуральных мышц в направлении от дистальных к проксимальным, от голеней до бедер и туловища для балансировки центра тяжести над опорным основанием (стопами).
Повреждение червя мозжечка нарушает нормальное упреждающее сокращение (через латеральный вестибуло-спинномозговой путь) медленных постуральных мышц, что приводит к потере равновесия в результате неспособности противостоять смещению центра тяжести, возникающему при движении разных частей тела. Поддержание позы.
Пациента просят произвести резкое разгибание в запястье с последующим коротким удержанием в разогнутом положении.
При регистрации ЭМГ видно, что сокращение сгибателей запястья начинается до завершения движения.
Обратите внимание на электрическую активность мышц-синергистов и антагонистов в положении «удержания».
ЭМГ-активность мышцы-антагониста более слабая, что показано на шкалах слева.
б) Мозжечок и высшие корковые функции. При позитронной эмиссионной томографии (ПЭТ) и фМРТ были получены данные о локальных изменениях кровотока и потребления кислорода. «Карты движений» построены на основе повторяющихся движений, таких как сжимание и разжимание кисти в кулак. Интересная особенность карт движений заключается в их очень малом размере и крайне медиальном расположении. До внедрения ПЭТ считали, что латеральное расширение задней доли мозжечка отвечает за праворукость. В настоящее время считают, что латеральное расширение может быть связано с когнитивными функциями (например, мышлением) и является отделом, взаимодействующим с латеральной предлобной (префронтальной) корой больших полушарий. Активация латерального расширения мозжечка становится максимальной при разговоре с доминированием одной из сторон, обусловленным возможными связями (с переключением в таламусе) с двигательным речевым центром лобной коры доминантного полушария. В некоторых случаях двигательный речевой центр осуществляет более строгий контроль в связи с тем, что латеральная доля мозжечка сильнее активизируется при функциональном обозначении объекта, например «копать» или «летать» вместо простого определения самого объекта — «лопата» или «самолет».
Мозжечковый когнитивно-аффективный синдром—недавно введенный собирательный термин, обозначающий функциональные корковые нарушения, возникающие вследствие внезапного тяжелого поражения мозжечка, например при тромбозе одной из трех пар мозжечковых артерий или неизбежного повреждения при удалении опухоли мозжечка. У таких пациентов развиваются когнитивные нарушения в виде снижения умственных способностей, невнимательности, появления грамматических ошибок в речи, нарушения пространственного чувства и частичной потери памяти. При сопутствующем поражении червя мозжечка возникают также аффективные (эмоциональные) нарушения, иногда в форме угнетения аффекта (притупления эмоциональных реакций) либо в виде аберрантного поведения. Когнитивно-аффективный синдром носит временный характер и может быть связан с уменьшением кровотока (при ПЭТ) в одной или более ассоциативных зонах, связанных с мозжечком корково-мосто-мозжечковыми волокнами. Помимо хорошо известных таламо-корковых путей к двигательной коре, мозжечок также может «управлять» таламическими нейронами, связанными с ассоциативными когнитивными и аффективными зонами.
Постурография — инструментальный метод исследования управления позой. Пациент стоит на платформе, а спонтанные колебания тела регистрируют датчики деформации под углами платформы. Информация от датчиков поступает в компьютер, что позволяет создать графическую запись колебаний тела в переднезаднем и боковом направлениях. Вначале исследование проводят с открытыми глазами, а затем — с закрытыми. Этот метод называют статической постурографией и используют для определения причины атаксии.
При динамической постурографии можно получить данные об эффекте внезапного смещения поддерживающей платформы на 4° кзади. В эту фазу исследования накладывают ЭМГ-электроды на икроножную (сгибатель стопы) и переднюю большеберцовую (разгибатель стопы) мышцы. В норме наблюдают тройную реакцию на смещение кзади:
(а) спинномозговое моносинаптическое рефлекторное сокращение икроножных мышц через 45 мс;
(б) полисинаптическое рефлекторное сокращение икроножных мышц через 95 мс;
(в) сокращение разгибателей стопы через гамма-петлю спустя 120 мс.
Восходящая дуга гамма-петли проходит в составе большеберцового и седалищного нервов в задний канатик через медиальный лемнисковый тракт к соматосенсорной коре; нисходящая дуга проходит через корково-спинномозговой путь, а также седалищный и малоберцовый нервы. Динамическая постурография позволяет дифференцировать широкое разнообразие заболеваний, поражающих центральную и периферическую нервную систему на различных уровнях.
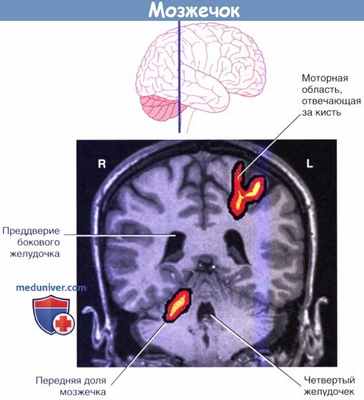
Изображение фМРТ-активности у добровольца, производящего повторяющиеся движения пальцев правой кисти.
У мозжечка нашли принципиально новую функцию
Эксперименты на мышах, проведенные американскими учеными, позволили обнаружить ранее неизвестную функцию мозжечка. Оказалось, что этот отдел мозга принимает участие в вознаграждении, или внутреннем подкреплении — механизме закрепления поведения путем положительных реакций на совершенное действие. Результаты работы опубликованы в журнале Nature.
Многочисленные наблюдения за животными и людьми с различными поражениями мозжечка давно привели ученых к выводу, что его функция состоит в тонкой настройке произвольных и непроизвольных движений, что обеспечивает четкую координацию, быструю реакцию, равновесие, осанку и ритмичное дыхание. Это подтверждало и наличие тесных нервных связей этого отдела мозга с корковыми и подкорковыми двигательными и сенсорными центрами.
Большинство нейронов мозжечка представлены гранулярными клетками, число которых превышает количество всех остальных нейронов головного мозга, при этом они занимают менее 10 процентов его объема. Из-за небольшого размера и плотного расположения изучать возбуждение отдельных гранулярных клеток весьма сложно. В сочетании с тем, что предназначение мозжечка казалось вполне понятным и интересовало ученых значительно меньше функций больших полушарий и подкорковых структур, это привело к тому, что клеточная физиология этого отдела мозга изучена недостаточно.
Чтобы разобраться, как гранулярные клетки мозжечка мышей функционируют в процессе совершения движений, сотрудники Стэнфордского университета воспользовались двухфотонной кальциевой визуализацией, позволяющей следить за возбуждением отдельных нейронов в реальном времени. В качестве унифицированной двигательной активности было выбрано нажатие передней лапой на рычаг в обмен на порцию сладкого раствора, подаваемого автоматически ко рту через небольшой промежуток времени (двухфотонная визуализация требует фиксации головы животного, поэтому более объемные движения при ее проведении нежелательны).
Выяснилось, что определенные нейроны, как и ожидалось, активируются при движении лапой. Неожиданным стало то, что во время ожидания награды (сладкого раствора) активируется другая группа нейронов, которая «выключается» при ее получении. Чтобы проверить, является ли эта активация реакцией на сенсорное восприятие сладости, ученые стали случайным образом не давать раствор в ответ на нажатие рычага и выяснили, что это активирует еще одну группу гранулярных клеток. Изменяя время между нажатием на рычаг и количество раствора и сопоставляя активацию разных групп нейронов с двигательной активностью, исследователи убедились, что активность нескольких групп гранулярных клеток связана не с движениями, а с реакцией на ожидаемую, полученную и не полученную награду.
Таким образом, функции мозжечка оказались шире, чем считалось ранее, и включают не только координацию движений, но и работу с вознаграждением. По словам руководителя работы Марка Вагнера (Mark Wagner), полученные результаты позволят полнее интегрировать мозжечок в целостную картину функционирования мозга.
Это не первый случай, когда классические представления о строении и функциях отделов нервной системы оказались ошибочными. Так, в 2016 году франко-британский научный коллектив доказал, что граница между симпатическим и парасимпатическим отделами автономной нервной системы была проведена неправильно.
Читайте также:
- Паукообразные ангиомы
- Анатомия: Эпикард. Эндокард.
- Кровоток при венозной гипертонии. Венозный тонус при венозной гипертонии
- Золотистый стафилококк. Staphylococcus aureus. Эпидемиология золотистого стафилококка.
- Рентгенограмма, МРТ, сцинтиграфия при энхондроме
Мозжечок — это функциональное ответвление главной оси «кора больших полушарий — спинной мозг». С одной стороны, в нём замыкается сенсорная обратная связь, то есть он получает копию афферентации (информации, передаваемой из спинного мозга в кору полушарий головного мозга), с другой стороны, сюда же поступает копия эфферентации (информации от коры полушарий к спинному мозгу) от двигательных центров. Говоря техническим языком, первая сигнализирует о текущем состоянии регулируемой переменной, а вторая даёт представление о требуемом конечном состоянии. Сопоставляя первое и второе, кора мозжечка может рассчитывать ошибку, о которой сообщает в двигательные центры. Так мозжечок непрерывно корректирует и преднамеренные, и автоматические движения. У низших позвоночных информация в мозжечок поступает также из акустической области, в которой регистрируются ощущения, относящиеся к равновесию, поставляемые ухом и боковой линией, а у некоторых даже от органа обоняния.
| Филогенетические отделы мозжечка | функция | соответствующие образования (у человека) |
|---|---|---|
| Архицеребеллум (вестибулоцеребеллум) | Взаимосвязь мозжечка с вестибулярными ядрами | Клочковая и узелковая дольки; ядро шатра |
| Палеоцеребеллум (спиноцеребеллум) | Взаимосвязь мозжечка со спинным мозгом | Червь мозжечка, околоклочковая долька; ядро шатра, пробковидное и круглое ядра |
| Неоцеребеллум (понтоцеребеллум) | Взаимосвязь мозжечка с корой больших полушарий головного мозга | Полушария мозжечка; зубчатое ядро |
Филогенетически наиболее древняя часть мозжечка (архицеребеллум) состоит из клочка (лат. flocculus) и узелка (лат. nodulus). Здесь преобладают вестибулярные входы (отсюда другое название этой части — вестибулоцеребеллум). В эволюционном плане структуры архицеребеллума возникают в классе круглоротых у миног, в виде поперечной пластинки, перекидывающейся через передний отделромбовидной ямки. У низших позвоночных (у рыб) архицеребеллум представлен парными ушковидными частями. В процессе эволюции (земноводных, пресмыкающихся, птиц и млекопитающих) отмечается уменьшение размеров структур древней части мозжечка. Архицеребеллум является важнейшей составляющей вестибулярного аппарата.
К «старым» структурам (палеоцеребеллум) у человека относятся также область червя (лат. vermis) в передней доле мозжечка, пирамиды, язычка червя и околоклочка (лат. paraflocculus). В палеоцеребеллум поступают сигналы в основном от спинного мозга. Структуры палеоцеребеллума появляются у рыб и представлены у других позвоночных.
Медиальные элементы мозжечка дают проекции к ядру шатра, а также к шаровидному и пробковидному ядрам, которые в свою очередь образуют связи главным образом со стволовыми двигательными центрами. Ядро Дейтерса — вестибулярный двигательный центр — тоже напрямую получает сигналы от червя и от флоккулонодулярной доли.
Повреждения архи- и палеоцеребеллума приводят в первую очередь к нарушениям равновесия, как и при патологии вестибулярного аппарата. У человека проявляется головокружением, тошнотой ирвотой. Типичны также глазодвигательные расстройства в виде нистагма. Больным трудно стоять и ходить, особенно в темноте (когда отсутствует зрительная коррекция положения в пространстве), для этого им приходится хвататься за что-нибудь руками; походка становится шатающейся, как будто в состоянии опьянения.
К латеральным элементам мозжечка (неоцеребеллум) идут сигналы преимущественно от коры полушарий головного мозга через ядра моста и нижней оливы. Клетки Пуркинье полушарий мозжечка дают проекции через латеральные зубчатые ядра к двигательным ядрам таламуса и далее к двигательным областям коры полушарий головного мозга. Через эти два входа полушария мозжечка получают информацию от корковых областей, активирующихся в фазу подготовки к движению, то есть участвующих в его «программировании». Структуры неоцеребеллума имеются только у млекопитающих. При этом у человека в связи с прямохождением, усовершенствованием движений руки они достигли наибольшего развития в сравнении с другими животными.
Таким образом, часть импульсов, возникших в коре мозга, достигает противоположного полушария мозжечка, принося информацию не о произведённом, а лишь о намечаемом к выполнению активном движении. Получив такую информацию, мозжечок моментально высылает импульсы, корригирующие произвольное движение главным образом путём погашения инерции и наиболее рациональной регуляции тонуса мышц агонистов и антагонистов. В результате обеспечивается чёткость и отточенность произвольных движений, устраняются какие-либо нецелесообразные компоненты.
Пластичность функций, двигательная адаптация и двигательное научение
Роль мозжечка в двигательной адаптации продемонстрирована экспериментально. Если нарушить зрение (например, поместив перед глазами призмы), вестибуло-окулярный рефлекс компенсаторного движения глаз при поворотах головы уже не будет соответствовать получаемой мозгом зрительной информации. Испытуемому в очках-призмах сначала очень трудно правильно перемещаться в окружающей среде, однако через несколько дней он приспосабливается к аномальной зрительной информации. При этом отмечены чёткие количественные изменения вестибуло-окулярного рефлекса, его долговременная адаптация. Опыты с разрушением нервных структур показали, что такая двигательная адаптация невозможна без участия мозжечка. Пластичность функций мозжечка и двигательное научение, определение их нейрональных механизмов было описано Девидом Марром и Джеймсом Альбусом.
Пластичность функции мозжечка ответственна также за двигательное научение и выработку стереотипных движений, таких как письмо, печатание на клавиатуре и др.
Хотя мозжечок и связан с корой головного мозга, его деятельность не контролируется сознанием.
Функции
Функции мозжечка сходны у различных биологических видов, включая человека. Это подтверждается их нарушением при повреждении мозжечка в эксперименте у животных и результатами клинических наблюдений при заболеваниях, поражающих мозжечок у человека. Мозжечок представляет собой мозговой центр, который имеет в высшей степени важное значение для координации и регуляции двигательной активности и поддержания позы. Мозжечок работает главным образом рефлекторно, поддерживая равновесие тела и его ориентацию в пространстве. Также он играет важную роль (особенно у млекопитающих) в локомоции (перемещении в пространстве).
Соответственно главными функциями мозжечка являются:
- координация движений
- регуляция равновесия
- регуляция мышечного тонуса
Проводящие пути
Мозжечок связан с другими отделами нервной системы многочисленными проводящими путями, которые проходят в ножках мозжечка. Различают афферентные (идущие к мозжечку) иэфферентные (идущие от мозжечка) пути. Эфферентные пути представлены только в верхних ножках.
Пути мозжечка не перекрещиваются вообще либо перекрещиваются дважды. Поэтому при половинном поражении самого мозжечка либо одностороннем поражении ножек мозжечка симптоматика поражения развивается на стороны поражения (гомолатерально).
Верхние ножки
Через верхние ножки мозжечка проходят эфферентные пути, за исключением афферентного пути Говерса.
- Передний спинально-мозжечковый путь (лат. tractus spino-cerebellaris anterior) (путь Говерса) — первый нейрон этого пути начинается от проприорецепторов мышц, суставов, сухожилий инадкостницы и находится в спинномозговом ганглие. Второй нейрон — клетки заднего рога спинного мозга, аксоны которого переходят на противоположную сторону и поднимаются вверх в передней части бокового столба, проходят продолговатый мозг, варолиев мост, затем вновь перекрещиваются и через верхние ножки поступают в кору полушарий мозжечка, а затем в зубчатое ядро.
- Зубчато-красный путь (лат. tractus dento-rubralis) — начинается от зубчатого ядра и проходят через верхние мозжечковые ножки. Эти пути осуществляют двойной перекрёст (перекрёст Вернекинга) и заканчиваются на красных ядрах (лат. nucleus rubrae). Аксоны нейронов красных ядер формируют руброспинальный путь (путь Монакова). После выхода из красного ядра этот путь вновь перекрещивается (перекрёст Фореля), спускается в стволе мозга, в составе бокового столба спинного мозга и достигает α- и γ-мотонейронов спинного мозга.
- Мозжечково-таламический путь (лат. tractus cerebello-thalamicus) — идёт к ядрам таламуса. Через них связывает мозжечок с экстрапирамидной системой и корой головного мозга.
- Мозжечково-ретикулярный путь (лат. tractus cerebello-reticularis) — связывает мозжечок с ретикулярной формацией, от которой в свою очередь начинается ретикулярно-спинальный путь.
- Мозжечково-вестибулярный путь (лат. tractus cerebello-vestibularis) — особый путь, так как в отличие от других проводящих путей, начинающихся в ядрах мозжечка, представляет собой аксоны клеток Пуркинье, направляющиеся к латеральному вестибулярному ядру Дейтерса (ядру преддверно-улиткового нерва).
Средние ножки
Через средние ножки мозжечка проходят афферентные пути, которые соединяют мозжечок с корой головного мозга.
- Лобно-мосто-мозжечковый путь (лат. tractus fronto-ponto-cerebellaris) — начинается от передних и средних лобных извилин, проходит через переднее бедро внутренней капсулы на противоположную сторону и переключается на клетках варолиевого моста, которые представляют собой второй нейрон данного пути. От них он поступает в контрлатеральную среднюю ножку мозжечка и заканчивается на клетках Пуркинье его полушарий.
- Височно-мосто-мозжечковый путь (лат. tractus temporo-ponto-cerebellaris) — начинается от клеток коры височных долей головного мозга. В остальном его ход схож с таковым лобно-мосто-мозжечкового пути.
- Затылочно-мосто-мозжечковый путь (лат. tractus occipito-ponto-cerebellaris) — начинается от клеток коры затылочной доли головного мозга. Передаёт в мозжечок зрительную информацию.
Нижние ножки
В нижних ножках мозжечка проходят афферентные проводящие пути идущий от спинного мозга и ствола головного мозга к коре мозжечка.
- Задний спинально-мозжечковый путь (лат. tractus spino-cerebellaris posterior) (путь Флексига) связывает мозжечок со спинным мозгом. Проводит импульсы от проприорецепторов мышц, суставов,сухожилий и надкостницы, которые достигают задних рогов спинного мозга в составе чувствительных волокон и задних корешков спинномозговых нервов. В задних рогах спинного мозга они переключаются на т. н. клетки Кларка, представляющие собой второй нейрон глубокой чувствительности. Аксоны клеток Кларка формируют путь Флексига. Они проходят в задней части бокового столба со своей стороны и в составе нижних ножек мозжечка достигают его коры.
- Оливо-мозжечковый путь (лат. tractus olivo-cerebellaris) — начинается в ядре нижней оливе с противоположной стороны и заканчивается на клетках Пуркинье коры мозжечка. Оливо-мозжечковый путь представлен лазящими волокнами. Ядро нижней оливы получают информацию непосредственно от коры головного мозга и таким образом проводит информацию от её премоторных зон, то есть областей ответственных за планирование движений.
- Вестибуло-мозжечковый путь (лат. tractus vestibulo-cerebellaris) — начинается от верхнего вестибулярного ядра Бехтерева (ядра преддверно-улиткового нерва) и через нижние ножки достигает коры мозжечка флоккуло-нодулярной области (архицеребеллума). Информация вестибуло-мозжечкового пути переключившись на клетках Пуркинье достигает ядра шатра(лат. nucleus fastigii).
- Ретикуло-мозжечковый путь (лат. tractus reticulo-cerebellaris) — начинается от ретикулярной формации ствола головного мозга, доходит до коры червя мозжечка. Соединяет мозжечок и базальные ганглии экстрапирамидной системы.