Медицинская реабилитация
7.2.1. Механизмы восстановления функций при повреждении периферических нервов и сплетений
Поврежденное нервное волокно не способно срастаться. Однако одновременно с процессом денервации начинаются восстановительные процессы, которые могут идти по трем направлениям.
(1) Регенерация нерва: проксимальная культя формирует аксональные выросты (наплывы аксоплазмы, или «колбы роста»), которые начинают продвигаться дистально и врастать в эндоневральные трубки (разумеется, лишь в тех случаях, когда последние сохранили свою целостность). Миелиновая оболочка вновь образующегося волокна формируется из тяжей леммоцитов. Скорость регенерации аксонов составляет приблизительно 1.5-2 мм в день [Buchtal F., Kuchl V., 1979]. Отдельные нервные проводники имеют различную способность к регенерации: среди периферических нервов особенно хорошо восстанавливается функция лучевого и мышечно-кожного нервов, а наихудшими регенеративными способностями обладают локтевой и малоберцовый нервы [Карчикян С.И., 1962; Weber R., 1996J. Для достижения хорошего восстановления растущие аксоны должны подключиться к дистальной культе нерва до того, как в ней наступят выраженные пери- и эндоневральные рубцовые сращения [Lefferet R., 1985]. В случаях формирования по ходу прорастающего волокна соединительнотканного рубца часть аксонов не распространяется в дистальном направлении, а беспорядочно отклоняется в стороны, формируя травматическую неврому.
При полном анатомическом повреждении нервного ствола через 2-3 недели после травмы на центральном конце образуется ампутационная неврома.
Регенерация нервного ствола может происходить гетерогенно: часть двигательных волокон врастает в оболочки чувствительных, а одноименные — в пучки, иннервирующие разноименные участки конечности [Гайдар Б.В.,1997].
(2) В случаях, когда в нервном стволе поражаются не все, а лишь часть нервных волокон, восстановление функции мышц возможно за счет разветвления сохранившихся аксонов и «захвата» ими тех мышечных волокон, которые иннервировались погибшими аксонами; при этом происходит укрупнение двигательных единиц мышцы. За счет этого механизма мышца может сохранять свою работоспособность в случаях утраты до 50% иннервировавших ее аксонов (а для мышц, не развивающих значительных усилий — даже до 90%), однако для полного завершения процесса компенсаторной иннервационной перестройки требуется около года [Weber R.,1996].
(3) В ряде случаев (обычно при травме нервного ствола типа ушиба) восстановление или улучшение функций связано с обратимостью некоторых патоморфологических процессов: с исчезновением реактивных воспалительных явлений, с рассасыванием мелких кровоизлияний и т.д. При нетяжелых травмах проводимость нерва, даже после полной ее потери, восстанавливается в течение первых дней или недель.
7.2.2. Факторы, определяющие прогноз восстановления
К основным факторам, определяющим при периферических невропатиях и плексопатиях скорость и степень спонтанного восстановления нарушенных функций (и, следовательно, объем и направленность лечебных вмешательств), относятся следующие:
— степень повреждения нервного проводника;
— уровень поражения;
— характер повреждающего агента.
7.2.2.1. Степень повреждения нервных проводников (при локальной травме)
Реабилитологи чаще всего определяют степень повреждения нерва по 3 категориям согласно классификации H.Seddon [1943]. Иногда используют также классификацию S.Sunderland [1990], выделяющего 5 степеней повреждения нервов; эта классификация основана на классификации H.Seddon, детализируя ее. Согласно классификации H.Seddon, все локальные повреждения нервных стволов делят, в зависимости от сохранности аксона и соединительнотканных структур, на три группы: (1) нейрапраксия; (2) аксонотмезис; (3) нейротмезис. (1) Нейрапраксия (neurapraxia, англ.) — это повреждение нерва, не приводящее к гибели аксона. Часто наблюдается при компрессии нерва (например, «ночной субботний паралич» вследствие компрессии лучевого нерва), при легкой травме нерва. Клинически характеризуется снижением вибрационной, проприоцептивной, иногда тактильной чувствительности. Болевая чувствительность страдает реже. Часто наблюдаются двигательные нарушения и парестезии. Блок проведения нервного импульса, наблюдающийся вследствие локального повреждения миелиновой оболочки, носит преходящий характер и регрессирует по мере восстановления миелина. Восстановление двигательных и чувствительных функций может продолжаться до 6 месяцев.
(2)Аксонотмезис (axonotmesis, англ.) — повреждение нерва, приводящее к гибели аксона при сохранности эпиневрия, периневрия, эндоневрия и шванновских клеток. Нередко наблюдается при закрытых переломах либо вывихах костей конечностей, а также при сдавлениях нервных стволов. Нарушаются двигательные, чувствительные и судомоторные функции нерва. Восстановление функций происходит за счет регенерации аксона. Скорость и степень восстановления зависит от уровня поражения, возраста (у молодых регенерация происходит быстрее) и общего состояния больного. В случаях, когда прорастание аксона происходит медленно, может произойти рубцевание эндоневральной трубки, в которую прорастает аксон, и восстановление не наступает. По этой же причине неблагоприятный прогноз имеется в случаях, когда дефект нервного ствола имеет значительную длину. При благоприятных условиях происходит постепенная невротизация дистального отдела поврежденного нерва, которая продолжается в течение многих месяцев, иногда год и более. Наблюдается восстановление утраченных функций, но не всегда полное.
О) Нейротмезис (neurotmesis, англ.) — разрыв нерва с пересечением аксона и соединительнотканных оболочек нерва. Из-за того, что повреждаются эндоневральные трубки, невозможным становится прорастание в них аксонов, регенерация аксонов приводит к образованию травматической невромы. Прогноз восстановления неблагоприятный. Данная классификация основана на микроскопических изменениях в нервном стволе. Макроскопически различить степень повреждения практически невозможно. Диагностика основана на динамическом клиническом и электрофизиологическом наблюдении. В связи с этим при закрытых травмах нервных стволов отечественные авторы нередко применяют иную классификацию, основанную на выделении следующих 4 форм поражения нервного ствола [Макаров А.Ю., Амелина О.А., 1998]: сотрясение, ушиб, сдавление, тракция. Сотрясение не сопровождается морфологическими изменениями в нерве, нарушения функции нерва кратковременны (не более 1-2 недель) и полностью обратимы. Ушиб нерва характеризуется возникновением мелких кровоизлияний, участков размозжения нервных волокон и пучков, что приводит к полному либо частичному нарушению проводимости, длительным и стойким выпадением функций. При сдавлении нерва степень нарушения проводимости зависит в первую очередь от длительности сдаачения: при своевременном удалении сдвливающих нерв субстратов (гематома, инородное тело, отломок кости и т.д.) может наблюдаться быстрое и полное восстановление проводимости, тогда как при длительном сдавлении в нервном стволе развиваются дегенеративные изменения. Отсутствие восстановления функции в течение 2-3 месяцев является критерием полного анатомического перерыва нерва. Тракция (например, тракдия ветвей плечевого сплетения при вправлении вывиха плеча) обычно сопровождается частичным нарушением функции, однако восстановление проводимости по нерву происходит достаточно длительно (в течение нескольких месяцев).
7.2.2.2. Уровень поражения
Чем проксимальнее поражение нервного ствола или сплетения (т.е. чем больше расстояние от места повреждения до периферических окончаний), тем хуже прогноз восстановления функции, поскольку тем больший срок требуется для прорастания нервного волокна и тем больше вероятность развития в эндоневральной трубке периферического отрезка нерва необратимых Рубцовых изменений [Lefferet R., 1985]. Так, например, по данным С.И.Карчикяна [1962], при ранениях седалищного нерва в верхней трети бедра первые движения стопы и пальцев появляются лишь спустя 15-20 месяцев и позже после наложения нервного шва, а при ранениях этого же нерва в нижней трети бедра — через 10-15 месяцев после оперативного вмешательства.
Наихудший прогноз отмечается при повреждениях на корешковом уровне, поскольку корешки спинальных нервов не регенерируют и не могут быть восстановлены хирургическим путем. Поражение корешков (обычно — отрыв корешка на шейном уровне), в отличие от поражения сплетения, характеризуется следующими признаками:
— интенсивная жгучая боль, иррадиирущая вдоль соответствующего дерматома;
— парализация паравертебральных мышц, иннервируемых задними ветвями спинальных нервов;
— паралич мышц лопатки вследствие нарушения функции коротких нервов плечевого пояса (крыловидная лопатка);
— синдром Горнера (при поражении С8-ты корешков);
— трофические нарушения и быстро прогрессирующая мышечная атрофия с грубыми вторичными контрактурами.
7.2.2.3. Характер повреждающего агента
Периферические невропатии и плексопатии могут иметь самую различную этиологию (таблица 7.2). В мирное время наиболее частой формой поражений периферических нервов являются туннельные невропатии, составляя около 30-40% от всех заболеваний периферической нервной системы. Туннельная невропатия — это локальное поражение нервного ствола, обусловленное его компрессией и ишемией в анатомических каналах (туннелях) или вследствие внешнего механического воздействия [Лейкин И.Б.,1998]. Предрасполагающие к развитию туннельных невропатий факторы включают генетически обусловленную узость естественных вместилищ нерва, приобретенную узость этих вместилищ вследствие отеков и гиперплазии соединительной ткани при различных заболеваниях (например, сахарном диабете, гипотиреозе, коллагенозах), длительное перенапряжение мышечно-связочного аппарата у лиц определенных профессий, последствия трав, мышечно-тонические и нейро-дистрофические нарушения при рефлекторных синдромах остеохондроза позвоночника, ятрогенные травмирующие воздействия (неправильное наложение гипсовой повязки, кровоостанавливающего жгута). Нарушение функции нерва происходит вследствие как демиелинизации, так и поражения аксона (ухудшение нейротрофического контроля в связи с недостаточностью аксонального транспорта).
Туннельные поражения нервов проявляются в первую очередь болью, чувствительными и вегетативными расстройствами. Двигательные нарушения развиваются лишь у одной трети больных и заключаются, как правило, в снижении мышечной силы, гипотрофии мышц, развитии контрактур. Прогноз восстановления функций при рано начатом лечении обычно благоприятный, однако это восстановление может происходить достаточно длительно, до нескольких месяцев. Кроме того, прогноз зависит от основного заболевания, на фоне которого развилась невропатия, от того, сохраняются ли профессиональные перегрузки конечности. В 30-40% случаев туннельные невропатии рецидивируют [Герман А.Г.и соавт., 1989].
На втором месте по частоте находятся травматические невропатии. Среди причин травматических невропатий прогностически наиболее благоприятными являются резаные ранения, при которых своевременное оперативное вмешательство обеспечивает хороший исход. Тракционные и огнестрельные травмы имеют худший прогноз, поскольку при них нередко измененными оказываются центральный отрезок нерва и нейрон спинномозговых центров, что существенно затрудняет регенерацию нерва. Разрушение нервного ствола на большом протяжении может наблюдаться также при электротравме, при химическом повреждении (случайное введение в нерв различных лекарственных веществ). Весьма неблагоприятно сопутствующее повреждению нерва нарушение кровообращения в конечности (кровотечение либо длительное наложение кровоостанавливающего жгута, тромбоз магистральной артерии), которое может приводить к развитию в мышцах, сухожилиях, суставных сумках, коже и подкожной клетчатке атрофирующего склерозирующего процесса с образованием контрактур. Препятствовать восстановлению движений могут также вторичные изменения в суставах и сухожилиях, которые развиваются вследствие растяжения связок и суставных сумок при пассивном свешивании конечностей в случае вялого паралича или пареза.
При невро- и плексопатиях, развившихся на фоне соматических заболеваний, вследствие иммунных, неопластических, инфекционных, токсических поражений и воздействий прогноз зависит от характера течения основного заболевания либо процесса.



7.2.3. Клинические и электрофизиоло-гические признаки восстановления нервных проводников
Определение степени восстановления функции нервных проводников основывается на данных комплексного клинико-электрофизиологического обследования, проводимого в динамике. Наиболее полное описание клинических закономерностей восстановления функции нервных проводников представлено в трудах, обобщающих опыт по лечению травматических невропатий, накопленный в период Великой Отечественной войны (Карчи кян С.И. Травматическое повреждение периферических нервов. — Л.:Медгиз, 1962; Аствацатуров М.И. Руководство по военной невропатологии. — Л., 1951; Опыт Советской медицины в Великой Отечественной войне,1952. — Т.20). Ниже мы рассмотрим закономерности восстановления функций после полного перерыва нерва в случае благоприятно протекающей регенерации либо после своевременно проведенного нейрохирургического вмешательства.
Наиболее ранними клиническими симптомами восстановления служат обычно изменения со стороны чувствительной сферы, которые задолго предшествуют признакам восстановления двигательной функции [Карчикян С.И., 1962]:
— парестезии, возникающие в зоне анестезии при давлении на область нерва тотчас ниже зоны повреждения, т.е. на область регенерирующих молодых аксонов;
— появление чувствительности к резкому сдавлению складки кожи в зоне анестезии;
— болезненность при давлении на нервный ствол дистальнее места повреждения с иррадиацией боли в дистальном направлении по ходу нерва; по мере прорастания аксона эта болезненность получается с уровней, отстоящих все более и более к периферии.
Восстановление чувствительности наступает раньше в более проксимальных отделах, начиная с краев центральной зоны анестезии. Вначале восстанавливается протопатическая (примитивная) болевая и температурная чувствительность: способность восприятия только резких болевых и температурных раздражений без точного распознавания качества и точной локализации наносимого раздражения. Поэтому болевые и температурные раздражения кожи вызывают ощущения, имеющие свойства гиперпатии (диффузные, трудно локализуемые, очень неприятные). Возможно, это связано с недостаточной миелинизацией новообразуюшихся регенерирующих волокон, что приводит к широкой иррадиации возбуждения на соседние волокна. Затем начинает восстанавливаться тактильная, и лишь потом — тонкая температурная чувствительность, мышечно-суставное чувство, стереогностическое чувство. По мере восстановления эпикритической (более тонкой) чувствительности начинают исчезать гиперпатические черты восприятия болевых и температурных раздражений.
Необходимо помнить, что сужение зоны расстройства чувствительности может происходить не только в результате начавшейся регенерации, но и за счет компенсаторных явлений (перекрытие ветвями соседних нервов); важно различать эти процессы.
К наиболее ранним признакам восстановления двигательной функции относятся некоторое повышение тонуса парализованных мышц, уменьшение атрофии. Затем, начиная с более проксимальных отделов, появляются активные мышечные сокращения. Через 5-6 месяцев после травмы нерва возникают активные движения, которые вначале отличаются слабостью, быстрой истощаемостью, неловкостью. Особенно долго длится восстановление мелких дифференцированных изолированных движений (например, в межфаланговых суставах). Позже всего восстанавливаются рефлексы, они нередко остаются утраченными даже при полном восстановлении чувствительности и двигательных функций. В целом поврежденный аксон при устранении причины, препятствующей аксональному росту, восстанавливается в сроки от 1.5-2 до 8-10 месяцев [Лобзин B.C. и соавт., 1988].
Даже при отсутствии регенерации частичное восстановление утраченных движений может происходить за счет компенсаторного сокращения мышц, иннервируемых сохранными нервами. С другой стороны, отсутствие восстановления движений может быть связано не с отсутствием регенерации нерва, а с сопутствующим повреждением сухожилий, мышц и суставов.
Среди электрофизиологических методов, используемых для динамического контроля за процессами восстановления нервной проводимости, в настоящее время применяют игольчатую и стимуляционную электромиографию (ЭМГ), а также метод вызванных потенциалов (глава 2 первого тома). Напомним, что частичное нарушение проводимости по нерву при регистрации стимуляционной ЭМГ характеризуется снижением скорости проведения возбуждения, снижением по амплитуде и частоте потенциалов действия нерва и мышцы, изменением структуры М-ответа; при регистрации игольчатой ЭМГ наблюдается изменение структуры потенциалов действия двигательных единиц соответствующих мышц. При демиелинизирующих процессах в большей степени снижается скорость проведения по нерву, тогда как при аксонопатиях наблюдается преимущественное снижение потенциала действия нерва и изменение М-ответа, а изменений скорости проведения может не наблюдаться. При полном перерыве нерва дистальный отрезок продолжает проводить импульсы до 5-6 суток. Затем наблюдается полное отсутствие электрической активности пораженных нервов и мышц. Спустя первые три недели обычно появляется спонтанная активность мышц в покое (денервационные потенциалы фибрилляций и положительные острые волны), регистрируемая с помощью игольчатых электродов. Первые признаки реиннервации после полной денервации мышцы выявляются при регистрации игольчатой ЭМГ в виде возникновения при попытке произвольного сокращения серии низковольтных полифазных потенциалов длительностью 5- 10 мс [Попов А.К., Шапкин В.И., 1997]. По мере реиннервации мышц наблюдается также появление полифазных двигательных единиц, увеличение их амплитуды и продолжительности (возникновение гигантских потенциалов двигательных единиц связывают с захватом сохранившимися аксонами дополнительных мышечных волокон). Потенциалы реиннервации иногда могут быть обнаружены за 2- 4 месяца до первых клинических признаков восстановления [Зенков J1.P., Ронкин М.А., 1991].
Наиболее ранние суждения о динамике восстановительных процессов могут быть получены с помощью регистрации вызванных потенциалов (ВП). Периферические ВП вызываются стимуляцией периферического нерва (магнитной или электрической) и регистрируются в виде волн над различными участками нерва. Через 7 и более дней после травмы сравнивают периферические ВП с теми, которые наблюдались сразу после травмы, либо с их значениями на непораженной контрлатеральной стороне. При этом ориентируются не только на амплитуду ВП, но и на площадь под волной ВП [Weber R.,1996]. При обратимом характере нарушений (нейрапраксия) спустя 7 и более дней после травмы на дистальном участке нерва продолжают регистрироваться сенсорные и моторные ВП, вызываемые стимуляцией периферического нерва ниже уровня поражения. При аксонотмезисе и нейротмезисе спустя этот срок наблюдается снижение амплитуды и изменение формы ВП дистальнее места повреждения, а по завершении валлеровского перерождения ВП на периферическом отрезке нерва не вызываются.
Электродиагностические методы позволяют отличить нейрапраксию от аксонотмезиса и нейротмезиса, но не позволяют различить между собой ак сонотмезис и нейротмезис; для этого необходимо использование метода магнитно-резонансной томографии [Weber R., 1996].
При плексопатиях метод ВII может помочь в дифференциальной диагностике пре- и постганглионарного поражений, что имеет важное значение при определении показаний к нейрохирургическому вмешательству. При постганглионарном поражении стволов сплетения дистальный конец теряет связь с телом клетки спинального ганглия, поэтому как сенсорный, так и двигательный потенциалы действия при раздражении периферического отрезка нерва отсутствуют в любой его точке ниже места повреждения. При преганглионарном поражении двигательные периферические ВП не вызываются при сохранности в тех же отрезках нерва чувствительных ВП (несмотря на анестезию в соответствующих зонах иннервации). Эго объясняется следующим: при преганглионарном поражении повреждению подвергается центральный отросток биполярной клетки, что нарушает передачу чувствительных импульсов к коре головного мозга и, соответственно, сопровождается анестезией. Однако периферический отрезок при этом не теряет связи с телом клетки спинального (чувствительного) ганглия, остается жизнеспособным и нормально проводит чувствительные импульсы. В связи с этим при преганглионарном повреждении сенсорный ПД регистрируется по всему ходу нервного волокна вплоть до уровня травмы. Ошибочные выводы, однако, могут быть сделаны при мультифокальной травме, когда имеется как пре-, так и постганглионарное корешковое повреждение; при этом сенсорные периферические ВП не вызываются, «маскируя» преганглионарное повреждение. Выявление преганглионарного поражения свидетельствует о крайне неблагоприятном прогнозе, поскольку, как уже указывалось, регенерация корешков невозможна, а хирургическое вмешательство недоступно.
Прогнозирование возможного спонтанного восстановления нарушенных функций определяет направленность и объем дальнейших реабилитационных мероприятий.

![]()
При большинстве нейродегенеративных заболеваний повреждается не только тело нейронов, но и аксоны, необходимые для передачи нервных импульсов. Механизмы, которые защищают нейроны от разрушения, одновременно ингибируют восстановление аксонов. Это большая проблема в разработке терапии заболеваний, связанных с потерей нейронов — от болезни Альцгеймера до глаукомы. Однако с помощью высокопроизводительного скрининга удалось найти киназы, ингибирование которых приводит и к сохранению нейрона, и к восстановлению аксонов. Хотя механизм этого эффекта до конца не понятен, обнаруженные киназы в будущем могут стать потенциальной мишенью для терапии нейродегенеративных заболеваний.
Развитие нервной системы и ее работа — постоянный баланс между клеточной смертью и выживанием, восстановлением и дегенерацией клеток и связей между ними. Нервная система состоит из множества взаимодействующих между собой нейронов. Соединения между ними, а также взаимодействие нейронов с рецепторами, мышцами и другими тканями и органами устанавливается не только во время эмбрионального развития, но и после рождения. Этот процесс включает в себя возникновение нейронов и их миграцию, появление синапсов, а также и постоянную работу над ошибками. Нейроны, которые неправильно взаимодействуют с соседними клетками или не смогли установить взаимодействие вовсе, и избыточные нейроны, которые были произведены нейрональными клетками-предшественниками «про запас», подлежат уничтожению путем апоптоза. Кроме того, существуют механизмы, которые редактируют только взаимодействие отростков нервных клеток (аксонов и дендритов), и не влияют на жизнеспособность клетки. Нарушение этого процесса может приводить к дефектам развития и нейродегенеративным заболеваниям — болезням Альцгеймера и Паркинсона, а также другим заболеваниям, связанным с потерей нейронов. Развитие и функционирование нервной системы регулируется в организме разными способами, один из которых — сигнальные каскады протеинкиназ.
Протеинкиназы — ферменты, которые присоединяют фосфатную группу к другим белкам (то есть фосфорилируют их). Фосфорилирование влияет на белки, например, активируя их способность к ферментативным реакциям или побуждая формировать белковые комплексы. Протеинкиназы нередко объединены в сигнальные каскады, состоящие из последовательных модулей. На вершине таких каскадов находятся протеинкиназы, реагирующие на внешние сигналы, в роли которых может выступать повреждение клетки, воздействие каких-либо веществ или избыток/недостаток питательных веществ. В ответ на сигнал протеинкиназа фосфорилируется, после чего она сама может фосфорилировать и активировать следующую киназу или киназы в каскаде. Классическим примером таких каскадов являются сигнальные пути митоген-активированных протеинкиназ (MAPK), которые, как правило, состоят из трех модулей (рис. 2).

Киназа киназы MAP-протеинкиназы DLK (Dual leucine zipper kinase — киназа с двойной лейциновой застежкой) — одна из ключевых протеинкиназ в нейронах млекопитающих. Она фосфорилирует протеинкиназы JNK (c-Jun N-terminal kinases), которые в свою очередь фосфорилируют огромное количество белков, связанных с транскрипцией генов, различными сигнальными путями, движением клеток, а также апоптозом. Таким образом, DLK участвует в огромном количестве процессов — порой противоположных друг другу. При развитии нервной системы DLK одновременно контролирует как образование аксонов и миграцию нейронов (за счет фосфорилирования белков, связанных с цитоскелетом), так и апоптоз лишних или нефункциональных нейронов (за счет активации соответствующих белков JNK-киназами). Кроме того, DLK участвует в дегенерации аксонов, но этот процесс не зависит от активности JNK-киназ. Во взрослом организме DLK запускает гибель нейронов в ответ на повреждение при ишемии, травме, отравлении нейротоксинами или нейродегенеративных заболеваниях (K. A. Fernandes et al., 2014. DLK-dependent signaling is important for somal but not axonal degeneration of retinal ganglion cells following axonal injury).
Повреждение аксонов сопутствует большому количеству патологических состояний, а также нормальному старению. Известно, что протеинкиназа DLK участвует в передаче сигнала от поврежденного участка аксона в тело нейрона, приводя к его апоптозу. Такой эффект хорошо изучен на примере ганглионарных клеток сетчатки глаза (см. картинку дня Конусы роста аксонов). Аксоны этих нейронов, объединяясь в зрительный нерв, передают зрительный импульс в мозг. При повреждении зрительного нерва сигнал о повреждении передается в тело нейронов на достаточно большое расстояние (длина зрительного нерва составляет примерно 30 мм).
Одной из стратегий по спасению ганглионарных клеток может быть подавление активности DLK. Действительно, при повреждении зрительного нерва не экспрессирующие DLK ганглионарные клетки выживают гораздо лучше (T. A. Watkins et al., 2013. DLK initiates a transcriptional program that couples apoptotic and regenerative responses to axonal injury). Но так как DLK также участвует в образовании аксонов, их регенерация невозможна, а восстановление функции нейронов становится все более маловероятным. Поэтому требуется поиск альтернативных методов, которые могли бы одновременно и сохранить поврежденный нейрон и обеспечить регенерацию аксонов.
Исследователи из Калифорнийского университета в Сан-Диего, которые ранее обнаружили роль DLK в регенерации аксонов (T. A. Watkins et al., 2013. DLK initiates a transcriptional program that couples apoptotic and regenerative responses to axonal injury), провели поиск новых киназ, которые важны как для сохранения нейрона при повреждении, так и для восстановления аксонов. Для поиска исследователи провели высокопроизводительный скрининг (high-throughput screening) 366 веществ, способных ингибировать 224 известные киназы. Некоторые из этих веществ подавляют активность не одной, а нескольких киназ, поэтому авторы провели скрининг двумя способами. Сначала они искали вещества, которые помогают выживать ганглионарным клеткам после повреждения. Так как каждую клетку нельзя повредить механически, к клеткам добавляли колхицин — вещество, мешающее работе микротрубочек цитоскелета, в том числе в аксонах. Такое химическое повреждение запускает те же механизмы, что и механическое. Во втором скрининге искали вещества, которые увеличивали рост отростков нейронов (аксонов и дендритов). В обоих случаях каждую из 366 молекул добавляли к культурам ганглионарных клеток, полученных из индуцированных плюрипотентных стволовых клеток человека, и наблюдали за исходом с помощью микроскопа.
Альтернативой такому подходу мог бы быть скрининг, в котором последовательно «выключали» гены с помощью малых интерферирующих РНК (siRNA, small interfering RNA). Но использование молекул решает две задачи. Во-первых, ингибиторы часто влияют на несколько киназ сразу, позволяя очертить круг из нескольких кандидатов, некоторые из которых могут оказаться неизвестными. Во-вторых, если вещество успешно снижает активность киназы, то оно сразу же становится кандидатом в лекарственные препараты или хотя бы в основу для нового лекарственного препарата. А это сохраняет огромное количество времени в последующих исследованиях.
По результатам скрининга вещества разделили на три группы: первая — неактивные вещества, вторая — вещества, увеличивающие выживаемость нейронов и (или) рост отростков нейронов, третья — вещества, уменьшающие эти параметры. При анализе полученных результатов удалось найти уже известные киназы. Ингибирование rho-зависимых киназ (Rho-associated protein kinase) ранее было связано с регенерацией аксонов ганглионарных клеток, а ингибирование циклинзависимых киназ — с их выживаемостью. Среди киназ, которые обеспечивали и то, и другое, выделялись три киназы, которые входят в подсемейство GCK-IV-киназ — MINK1, MAP4K4 и TNIK (рис. 3). Интересно, что эти киназы участвуют в активации DLK в нейронах в ответ на стресс (M. Larhammar et al., 2017. The Ste20 Family Kinases MAP4K4, MINK1, and TNIK Converge to Regulate Stress-Induced JNK Signaling in Neurons).

Роль киназ-кандидатов, безусловно, проверили с помощью альтернативных методов. Снижение их активности ранее известными ингибиторами приводило к увеличению выживаемости ганглионарных клеток и росту аксонов в клеточных культурах. Авторы также смогли изучить последствия того, что будет, если нарушить работу этих киназ в ганглионарных клетках, полученных из сетчатки мышей: снижение активности одновременно трех киназ увеличивало их выживаемость в культуре. Кроме того, нарушение работы киназ защищало нейроны от различных видов повреждений (окислительного стресса, повреждений микротрубочек, повреждения ДНК). После таких результатов настало время исследовать, как протеинкиназы GCK-IV действуют в живом организме.
Исследователям достаточно было изучить, как «выключение» активности киназ влияет на ганглионарные клетки. Поэтому они воспользовались аденоассоциированными вирусами, несущими направляющую РНК (guide RNA) против трех киназ, которые вводили напрямую в глаза трансгенных мышей, экспрессирующих нуклеазу Cas9, или контрольных мышей. Через 14 дней удалось достичь «выключения» целевых генов.
Далее у мышей разрушали зрительный нерв (подробности этой методики описаны в статье Z. Tang et al., 2011. An Optic Nerve Crush Injury Murine Model to Study Retinal Ganglion Cell Survival). Эта процедура имитирует то, что происходит в сетчатке пациентов с глаукомой или травмой зрительного нерва: после нарушения целостности аксонов сигнал о повреждении передается в тело ганглионарных клеток, и они отмирают путем апоптоза. Мышам с модифицированными нейронами и контрольным мышам проводили эту процедуру (или имитировали ее), а затем изучали, как выживают ганглионарные клетки. Гипотеза подтвердилась: через две и через десять недель после операции у мышей с нефункциональными протеинкиназами GCK-IV в сетчатке оставалось больше ганглионарных клеток, чем у контрольных животных. Такой же эффект наблюдался при «выключении» уже известной киназы DLK.
А что с аксонами? Авторы повторили описанную выше процедуру, включив в эксперимент вирусы с направляющей РНК против киназы DLK. Через две недели у мышей разрушали зрительный нерв, а через три недели изучали вновь образованные аксоны. У контрольных мышей наблюдали новые, но редкие отростки ганглионарных клеток. У мышей с неактивной киназой DLK, как и предполагалось, аксоны не восстанавливались. В случае неактивных GCK-IV-киназ новые аксоны появлялись, но в недостаточном количестве. Тогда авторы пошли еще на одну уловку — «выключение» гена PTEN. Этот способ помогает восстановить синтез белка в поврежденных нейронах (K. K. Park et al., 2008. Promoting Axon Regeneration in the Adult CNS by Modulation of the PTEN/mTOR Pathway) — еще одна проблема, которую нужно преодолеть для регенерации аксонов. Снижение уровня PTEN в ганглионарных клетках без DLK не приводит к восстановлению аксонов, зато в клетках, где отсутствовали GCK-IV-киназы, аксоны восстанавливались значительно лучше.

![]()
Пока авторы не могут объяснить, за счет чего ингибиторы GCK-IV-киназ одновременно увеличивают выживаемость самих нейронов и их аксонов. Одна из вероятных гипотез состоит в том, что GCK-IV-киназы участвуют в активации DLK. При их «выключении» DLK активируется, но не настолько, чтобы запустить клеточную смерть. В пользу этой гипотезы свидетельствует лишь частичная регенерация аксонов после повреждения зрительного нерва. Вторая гипотеза предполагает, что GCK-IV-киназы участвуют в ответе на повреждение аксонов независимо от DLK. Например, киназа MAP4K4 активируется в ответ на повреждение кардиомиоцитов при окислительном стрессе. «Выключение» этой киназы увеличивает выживаемость и улучшает функцию клеток в этих условиях (L. R. Fiedler et al., 2019. MAP4K4 Inhibition Promotes Survival of Human Stem Cell-Derived Cardiomyocytes and Reduces Infarct Size In Vivo). При этом в кардиомиоцитах DLK отсутствует.
Тем не менее, наличие еще одной мишени для разработки препаратов, помогающих восстановить утраченные или поврежденные нейроны, не может не радовать. Дальнейшее изучение того, как работают GCK-IV-киназы в нейронах при различных условиях, поможет в этом процессе.
Источник: Amit K. Patel, Risa M. Broyer, Cassidy D. Lee, Tianlun Lu, Mikaela J. Louie, Anna La Torre, Hassan Al-Ali, Mai T. Vu, Katherine L. Mitchell, Karl J. Wahlin, Cynthia A. Berlinicke, Vinod Jaskula-Ranga, Yang Hu, Xin Duan, Santiago Vilar, John L. Bixby, Robert N. Weinreb, Vance P. Lemmon, Donald J. Zack, and Derek S. Welsbie. Inhibition of GCK-IV kinases dissociates cell death and axon regeneration in CNS neurons // PNAS. 2020. DOI: 10.1073/pnas.2004683117.
Екатерина Грачева
Р.Ф. МАСГУТОВ, А.А. РИЗВАНОВ, А.А. БОГОВ (мл.), А.Р. ГАЛЛЯМОВ, А.П. КИЯСОВ, А.А. БОГОВ
Республиканская клиническая больница МЗ РТ, г. Казань
Казанский (Приволжский) федеральный университет
Масгутов Руслан Фаридович
старший научный сотрудник Травматологического центра
420064, г. Казань, ул. Оренбургский Тракт, д. 138, тел. 8-950-314-02-93, е-mail: [email protected]
В обзоре освещены современные методы терапии травмы периферических нервов в сочетании с классическими хирургическими способами лечения данной патологии. Представлены потенциальные стимуляторы посттравматической регенерации периферических нервов, ключевыми из которых являются сосудистый эндотелиальный фактор роста и основной фактор роста фибробластов, играющих роль узловых молекул в патофизиологии нейрорегенерации.
Ключевые слова: травма периферического нерва, нейротрофические факторы.
R.F. MASGUTOV, A.A. RIZVANOV, A.A. BOGOV (JR.), A.R. GALLYAMOV, A.P. KIYASOV, A.A. BOGOV
Republican Clinical Hospital of the Ministry of Health of the Republic of Tatarstan, Kazan
Kazan (Volga region) Federal University
Current trends for treatment of peripheral nerves injuries
The review describes current treatment methods of peripheral nerve injuries in combination with classical surgical treatments of this pathology. Are presented potential stimulators of posttraumatic regeneration of peripheral nerves, where the key stimulators are vascular endothelial growth factor and basic fibroblast growth factor, which are basic molecules in the pathophysiology of neuroregeneration.
Key words: peripheral nerve injury, neurotrophic factors
Повреждения периферических нервов остаются одной из актуальных проблем современной травматологии и составляют 3-10% от всех травм опорно-двигательного аппарата [1]. Между тем травмы и заболевания периферической нервной системы приводят к стойкой утрате трудоспособности у 60-63% пациентов, более того, почти 80% этой группы составляют лица молодого трудоспособного возраста [2]. Повреждения периферических нервов оказывают существенное влияние на качество жизни в связи с потерей функции и повышения риска развития вторичной инвалидности от падений, переломов и других травм.
В представленном обзоре литературе дан анализ исследований последних лет, которые открывают новую страницу в понимании патофизиологии посттравматической регенерации периферического нерва и направляют как специалистов фундаментальных направлений, так и практикующих врачей на изучение и возможности клинического применения ростовых факторов, которые непосредственно принимают участие в восстановлении периферического нерва после его травмы.
Патогенез травмы периферического нерва
При повреждении нерва основным звеном патогенеза является деструкция нейронов.
Нейроны, формирующиеся в процессе развития, соединены в сложные коммуникационные сети для передачи информации от периферических рецепторов сенсорных нейронов в ЦНС (головного и спинного мозга), а также для передачи команд из центральной нервной системы на эффекторные органы, такие как скелетные мышцы, иннервируемые мотонейронами.
Периферический нерв состоит из аксона нейрона, Шванновских клеток, фибробластов, а также элементов кровоснабжения нерва. Соединительная ткань известная как эндоневрий окружает периферические аксоны. В периферических нервах аксоны сгруппированы в фасцикулы, окруженные соединительной тканью. Ключевой клеткой для периферических нервов является Шванновская клетка. Шванновские клетки образуют осевые цилиндры, их мембрана богата липидами, которые покрывают аксон и называется миелиновой оболочкой. Шванновские клетки с миелиновой оболочкой аксонов поддерживают и направляют аксоны во время регенерации нерва после его травмы. Тело нервной клетки является местом синтеза практически всех белков и органелл в клетке. Сложный процесс, известный как антероградный транспорт, непрерывно транспортирует вещества из тела клетки нейрона через аксон к его периферическому концу — в синапс. Эти транспортируемые вещества включают нейротрансмиттеры, которые обеспечивают связь между нейроном и тканями органа по узкой синаптической щели или, как в случае иннервации двигательных нейронов мышц, нервно-мышечном синапсе. С другой стороны, клетки-мишени, например, миоциты, экспрессируют вещества, которые действуют как нейротрофические факторы. Некоторые из этих веществ упаковываются и передаются путем ретроградного транспорта от синапса через аксон к телу нейрона. Таким образом, нейрон и его клетка-мишень постоянно информированы о статусе связи между ними. Было высказано предположение, что информация от клеток-мишеней принимает форму факторов, которые поддерживают существующие соединения нейронов и способствуют регенерации поврежденных нервных клеток. Несмотря на регенерацию, обширные травмы периферического нерва могут привести к параличам всей конечности или дистальных отделов конечностей. Прогноз восстановления при травмах периферических нервов лучше там, где повреждение Шванновских клеток минимально [3].
Нейротрофические факторы в механизме регенерации периферического нерва
За полвека исследований выявлено, что нейротрофические факторы способствуют выживанию и процессу роста нейронов. Восстановление нейрона критически зависит от количества нейротрофического фактора. После травмы нервов, периферическая нервная система млекопитающих реагирует путем экспрессии нейротрофических факторов, которые доступны из аутокринных или паракринных источников. Известны три основные группы нейротрофических факторов: 1) нейротрофины; 2) нейротрофические факторы семейства GDNF (англ. Glial cell line-derived neurotrophic factor) и 3) нейротрофические цитокины [4].
Повреждение периферического нерва вызывает ретроградную дегенерацию спинальных ганглиев, что сказывается преимущественно на кожных нейронах малого диаметра. Результаты исследований показывают, что кожные нейроны более чувствительны к повреждениям периферического нерва, чем мышечные, но их регенеративные способности не отличаются друг от друга [5].
Выживаемость нейронов после аксотомии является необходимым условием для регенерации, которой способствуют множество трофических факторов из нескольких источников, в том числе нейротрофины, нейропоэтические цитокины, инсулиноподобный фактор роста (англ. Insulin Growth Factor, IGF), факторы из глиальных клеток — семейства GDNF и др.
Так, после перерезки нерва увеличивается количество нейронов, экспрессирующих м-РНК мозгового нейротрофического фактора (англ. Brain Derived Neurotrophic Factor [6]. Выработка м-РНК BDNF в ответ на травму нерва показана не только в нейронах, но и в клетках-сателлитах [7].
После лигирования спинномозговых нервов уровень м-РНК фактора роста нервов (англ. Nerve Growth Factor, NGF) в спинальных ганглиях возрастает в 4 раза и поддерживается в течение 3 недель [8]. При этом подъем уровня м-РНК BDNF длится не более трех суток и менее выражен по сравнению с NGF.
Трансформирующий фактор роста β (TGFβ) служит митогеном для шванновских клеток и нейротрофическим фактором для некоторых дифференцирующихся нейронов. В интактных спинальных ганглиях TGFβ и его рецептор экспрессируются преимущественно в малых нейронах и в клетках-сателлитах, окружающих средние и большие нейроны [7]. После повреждения седалищного нерва крысы в эксперименте в клетках-сателлитах увеличивается экспрессия TGFβ, что сопровождается возрастанием уровня экспрессии его рецепторов практически во всех субпопуляциях нейронов [7].
In vitro GDNF стимулирует рост аксонов чувствительных нейронов, преимущественно малых, не экспрессирующих рецепторы нейротрофинов и выявляемых при помощи изолектин-B4 [9]. Эти авторы установили, что в той же культуре GDNF поддерживает выживание чувствительных нейронов, но не выявили их принадлежности к конкретным субпопуляциям (табл. 1).
Таблица 1.
Влияние нейротрофических факторов на выживание чувствительных нейронов в постнатальном периоде
|
Нейротрофический фактор |
Наличие (+) или отсутствие (–) эффекта |
Авторы |
|
NGF |
+ – |
Horie, Akahori, 1994;Lindsay, 1996 |
|
BDNF |
– + |
Lindsay, 1996;Acheson et al., 1995 |
|
NT-3 |
+ |
Acheson et al., 1995 |
|
GDNF |
+ |
Leclere et al., 1997 |
Установлено, что часть нейронов, экспрессирующих trkA и реагирующих, таким образом, на NGF, в конце эмбрионального и в постнатальном периоде перепрограммируется на действие другого нейротрофического фактора, а именно GDNF. В нейронах этой субпопуляции начинают экспрессироваться оба компонента рецептора GDNF — GFRa и тирозинкиназа c-ret [10]. Происходит ли подобное перепрограммирование при регенерации чувствительных нейронов, остается неясным.
Регенераторный рост центральных отростков нейронов различного фенотипа поддерживают конкретные нейротрофические факторы. Так, рост аксонов, экспрессирующих NF200 и принадлежащих субпопуляции больших проприоцептивных нейронов, стимулирует только один представитель семейства нейротрофинов — нейротрофин-3 (NT-3) [11] (табл. 2). Из тех же нейротрофинов только NGF поддерживает рост аксонов пептидергических ноцицептивных нейронов. И только один нейротрофический фактор является универсальным стимулятором роста аксонов больших, малых пептидергических и малых непептидергических нейронов. Этим фактором является GDNF [11].
Таблица 2.
Экспрессия нейротрофинов и их рецепторов нейронами спинальных ганглиев (по результатам экспериментов на нокаутных мышах [12]
|
Нейротрофин/ Рецептор |
Численность популяции нейронов в % |
Модальность |
|
NGF |
80 |
Ноцицепторы, терморецепторы, низкопороговые механорецепторы |
|
trkA |
80 |
Ноцицепторы, терморецепторы, низкопороговые механорецепторы |
|
trkB |
30 |
Механорецепторы (Майсснера) |
|
NT-3 |
60 |
Проприорецепторы, рецепторы волос и механорецепторы |
|
trkC |
20-30 |
Проприорецепторы |
После перерезки седалищного нерва усиливается экспрессия рецептора GDNF, но количество нейронов, реагирующих на этот нейротрофический фактор, не изменяется [13]. При этом объем популяции нейронов, содержащих trkA и отвечающих на действие NGF, после перерезки нерва уменьшается. Эти данные указывают на различный характер регуляции активности генов рецепторов нейротрофических факторов в субпопуляциях нейронов спинальных ганглиев в ответ на аксотомию.
Для судьбы отдельных нейронов важно присутствие нейротрофических факторов, которые способны активировать антиапоптозные внутриклеточные пути [14]. В списке факторов, влияющих на регенерацию нервных волокон, важное место занимают цитокины. Имеются данные о нейропротекторном действии интерлейкина-1b в отношении нейронов коры, которое может реализоваться через влияние этого цитокина на выработку нейротрофических факторов, например, NGF [15]. С другой стороны, in vitro показано, что интерлейкин-1b не влияет на выживание нейронов спинальных ганглиев, но ускоряет рост их отростков [16] и вместе с оксидом азота поддерживает в пептидергических нейронах экспрессию циклооксигеназы-2, что усиливает секрецию вещества «Р» [17]. Высказано предположение, что данный цитокин стимулирует секрецию нейротрофических факторов ненервными (Шванновскими) клетками [16], расположенными в потенциальном пространстве роста аксонов.
Представление об активации интерлейкином-1b регенерации нервных проводников основано на данных о стимулирующем влиянии продуцируемого макрофагами интерлейкина-1 на образование NGF шванновскими клетками in vitro и in vivo. Шванновские клетки in vitro и в регенерирующем нерве сами синтезируют интерлейкины и экспрессируют их рецепторы, что свидетельствует о возможной аутокринной регуляции. Нейроны также способны синтезировать цитокины. У интактных крыс до 80% нейронов спинального ганглия экспрессируют интерлейкин-1b [18]. Это преимущественно большие и средние нейроны. Данный цитокин экспрессируется в незначительной части малых пептидергических нейронов. Присутствие рецепторов интерлейкина-1 выявлено в большинстве нейронов спинального ганглия, а также в окружающих нейроны клетках-сателлитах [18], что свидетельствует об участии интерлейкина-1b в ауто/паракринной регуляции функции чувствительных нейронов.
Травмированные нейроны должны перейти от режима передачи к режиму роста и начать продуцировать белки, такие как, например, GAP-43 (англ. Growth Associated Protein), тубулин и актин, а также другие нейропептиды и цитокины, которые имеют потенциал для стимуляции аксональной регенерации. Аксоны должны достигать дистального отрезка нерва при условии, когда поддержка их роста осуществляется в достаточной степени. Шванновские клетки в дистальном отрезке нерва проходят через фенотипические изменения, которые должны быть благоприятны для аксональной регенерации. Шванновские клетки играют незаменимую роль в содействии регенерации за счет увеличения ими синтеза поверхностных молекул клеточной адгезии, таких как N-CAM (англ. Neural Cell Adhesion Molecule), Ng-CAM/L1 (англ. Neuron Glia Cell Adhesion Molecule), N-кадгерин и L2/HNK-1, путем изменения базальной мембраны, которая содержит многие белки внеклеточного матрикса, таких как ламинин, фибронектин и тенасцин. Аксональной регенерации могут способствовать факторы, которые повышают потенциал роста аксонов нейронов и оптимизируют регенерацию дистального конца нерва в сочетании с его оперативным лечением [19].
Основные методы лечения повреждения нервов являются: нейрорафия, нейрорафия с последующей дистракцией нерва, аутонервная пластика, эндопротезирование биодеградирующими материалами (например: neuragen® nerve guide).
Перспективными потенциальными стимуляторами регенерации периферических нервов являются сосудистый эндотелиальный фактор роста (англ. Vascular Endothelial Growth Factor, VEGF), основной фактор роста фибробластов (англ. basic Fibroblast Growth Factor, bFGF или FGF2) и NGF.
Эффект NGF иллюстрирует исследование по трансплантации в поврежденный седалищный нерв крысы микрокапсул, содержащих NGF в чистом виде. В группе с NGF количество восстановленных аксонов было большим, расположение нервных волокон было более упорядоченным, проводимость нерва восстанавливалась в разы быстрее, чем в контрольной группе. Также в сравнении с контрольной группой животных было отмечено, что в послеоперационном периоде раны заживали значительно быстрее [20].
Основной фактор роста фибробластов считается мощным средством для улучшения восстановления после повреждения нейронов, однако экзогенно применяемый bFGF неэффективен из-за кратковременного эффекта. Для усиления эффекта в естественных условиях разработана новая система доставки потенциальных стимуляторов путем внедрения bFGF в желатин гидрогель, который медленно биодеградирует. В этом исследовании эффекты bFGF-гидрогеля при травматическом повреждении лицевого нерва были исследованы на морских свинках. Лицевой нерв был обнажен и зажат в вертикальной части с использованием микрощипцов. Животные были подвергнуты одной из следующих трех процедур: группа А — контроль, без введения bFGF; группа В, одноразовое введение bFGF в нерв; группа С, применение bFGF-гидрогеля. Шесть недель спустя функции лицевого нерва были оценены тремя тестами: наблюдения за мимикой, электрофизиологические тестирования и гистологические исследования. Результаты показали, что одноразовое применение bFGF не влияло посттравматическое восстановление нерва. В группе с применением гидрогеля достигнуты лучшие результаты во всех тестах [21].
Местное применение VEGF индуцирует, по крайней мере, два события: пролиферацию Шванновских клеток и образование новых сосудов, которые играют важную роль в процессе регенерации нервов. Однако результаты показывают, что эффект от предварительной обработки факторами роста является локальным и ограниченным и не влияет на синтез нейропептидов [22].
Преодоление диастаза периферического с помощью биодеградирующией трубки из политетрафторэтилена (ПТФЭ) является относительно новым, но хорошо зарекомендовавшим хирургическим подходом для восстановления дефектов нервных стволов до 4 см между его концами на разных уровнях верхней конечности [23].
Восстановление периферического нерва с помощью трубки из ПТФЭ и введением факторов роста фибробластов также подтверждается в другом исследовании. В данном исследовании bFGF был помещен в эндопротез для седалищного нерва крысы. Кроме того, в послеоперационном периоде, животные системно получали 1 мг/кг/сут FK506 (такролимус) — мощный иммунодепрессант с нейротрофическими свойствами. Регенерация нерва оценивалась с помощью стандартных двигательных (индекс седалищного нерва) тестов и тестов на чувствительность, начиная со 2-й недели после операции. Животные, которые получали bFGF и FK506, показали значительно более быстрое восстановление после травмы, чем у контрольной группы [24].
После повреждения периферического нерва Шванновские клетки начинают экспрессировать bFGF, что нехарактерно для интактного нерва. Данный фактор рассматривается в качестве узловой молекулы в патофизиологии травмы периферических нервов [25].
В исследовании по восстановлению лицевого нерва после его перерезки и дальнейшего сшивания на крысах было показано, что у животных с введением bFGF восстановление нерва произошло раньше, а количество миелинизированных волокон было большим, чем у животных без введения bFGF [26].
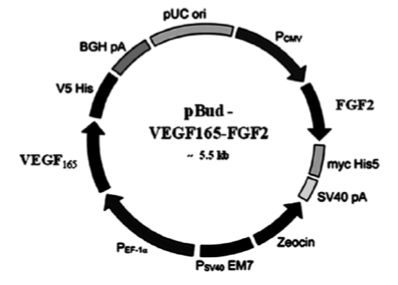
Кроме того, на модели преодоления диастаза седалищного нерва крысы при помощи аутонервной вставки показано, что введение мультицистронной плазмиды pBUD-VEGF-FGF2 (Рис 1), экспрессирующей клонированные гены VEGF и bFGF человека, в центральный и периферический отрезки нерва, а также непосредственно в аутонервную вставку стимулирует реваскуляризацию и регенерацию периферического нерва. В эксперименте с аутонервной вставкой введение pBUD-VEGF-FGF2 в область повреждения нерва приводит к двукратному увеличению количества S100-иммунопозитивных (Шванновских) клеток в прилежащих к диастазу участке периферического отрезка нерва [27, 28].
Рисунок 1.
Плазмида, содержащая сосудистый эндотелиальный фактор роста (VEGF) и основной фактор роста фибробластов (FGF2).
Масгутов Р.Ф. и др., Клеточная трансплантология и тканевая инженерия, 2011
Заключение
Таким образом, сочетание классических хирургических способов восстановления нервных стволов и методов прямой терапии ростовыми факторами ускоряет аксональный рост, стимулирует реваскуляризацию нерва, что подтверждается многочисленными экспериментальными исследованиями и, в итоге, улучшает результаты посттравматического восстановления функции реиннервации поврежденного органа или ткани.
Благодарность
Выполнение данного научного исследования финансируется за счет темы государственного автономного учреждения здравоохранения «Республиканская клиническая больница Министерства здравоохранения Республики Татарстан» «Усовершенствование и разработка новых методов лечения у больных с повреждением плечевого сплетения и периферических нервов».
ЛИТЕРАТУРА
1. Берснев В.П. Хирургия позвоночника, спинного мозга и периферических нервов / Е.А. Давыдов, Е.Н. Кондаков // Специальная литература. — СПб, 1998. — С. 368.
2. Кубицкий А.А. Хирургическое лечение повреждений периферических нервов верхней конечности методами тракционного удлинения и аутонервной пластики: автореф. дис. … канд. мед. наук / А.А. Кубицкий. — Казань, 2002. — 24 с.
3. Massing M.W., Robinson G.A., Marx C.E., Alzate O., Madison R.D. Frontiers in Neuroscience. Alzate O., editor // Source Neuroproteomics. — Boca Raton (FL): CRC Press; 2010. — Chapter 15.
4. Boyd J.G., Gordon T. Neurotrophic factors and their receptors in axonal regeneration and functional recovery after peripheral nerve injury // Mol Neurobiol. — 2003 Jun. — Vol. 27, № 3. — Р. 277-324.
5. Welin D., Novikova L.N., Wiberg M., Kellerth J.O., Novikov L.N. Survival and regeneration of cutaneous and muscular afferent neurons after peripheral nerve injury in adult rats // Exp Brain Res. — 2008 Mar. — Vol. 186, № 2. — Р. 315-23.
6. Zhou X.F. et al. Injured primary sensory neurons switch phenotype for brain-derived neurotrophic factor in the rat // Neuroscience. — 1999. — Vol. 92, № 3. — P. 841 853.
7. Zhou X.F. et al. Satellite-cell-derived nerve growth factor and neurotrophin-3 are involved in noradrenergic sprouting in the dorsal root ganglia following peripheral nerve injury in the rat // Eur. J. Neurosci. — 1999. — Vol. 11, № 5. — P. 1711 1722.
8. Shen H., Chung J., Chung K. Expression of neurotrophin mRNAs in the dorsal root ganglion after spinal nerve injury // Brain Res. Mol. — 1999. — Vol. 64. — P. 186 192.
9. Leclere P. et al. Effects of glial cell line-derived neurotrophic factor on axonal growth and apoptosis in adult mammalian sensory neurons in vitro. Neuroscience. — 1997. — Vol. 5. — P. 545 558.
10. Molliver D.C. et al. IB4 binding DRG neurons switch from NGF to GDNF dependence in early postnatal life // Neuron. — 1997. — Vol. 19, № 4. — P. 4849 4861.
11. Ramer M., Priestley J., McMahon S. Functional regeneration of sensory axons into adult spinal cord // Nature. — 2000. — Vol. 403. — P. 312 316.
12. Farinas I., Cano-Jaimeza M., Bellmunta E., Soriano M. Regulation of neurogenesis by neurotrophins in developing spinal sensory ganglia // Brain Research Bulletin. — 2002. — Vol. 57, № 6. — P. 809 816.
13. Kashiba H., Hyon B., Senba E. Glial cell line-derived neurotrophic factor and nerve growth factor receptor mRNAs are expressed in distinct subgroups of dorsal root ganglion neurons and are differentially regulated by peripheral axotomy in the rat // Neuroscience Letters. — 1998. — Vol. 4. — P. 107 110.
14. Mattson M.P., Lindvall O. Neurotrophic factor and cytokine signaling in the aging brain // Greenwich. JAI Press. — 1997. — P. 299 345.
15. Strijbos P., Rothwell N. Interleukin-1 beta attenuates excitatory amino acid-induced neurodegeneration in vitro: Involvement of nerve growth factor // J. Neurosci. — 1995. — Vol. 15, № 5. — P. 3468 3474.
16. Horie H., Sakai I., Akahori Y., Kadoya T. IL-1 beta enhances neurite regeneration from transected-nerve terminals of adult rat DRG // Neuroreport. — 1997. — Vol. 8, № 8. — P. 1955-1959.
17. Morioka N. et al. Nitric oxide synergistically potentiates interleukin-1 beta-induced increase of cyclooxygenase-2 mRNA levels, resulting in the facilitation of substance P release from primary afferent neurons: involvement of cGMP-independent mechanisms // Neuropharmacology. — 2002. — Vol. 43, № 5 10. — P. 868 876.
18. Copray J.C. et al. Expression of interleukin-1 beta in rat dorsal root ganglia // Neuroimmunology. — 2001. — Vol. 118, № 8. — P. 203 211.
19. Fu S.Y., Gordon T. The cellular and molecular basis of peripheral nerve regeneration // Mol Neurobiol. — 1997. — Vol. 1-2. 67-116.
20. Song M., Chen S.Z., Han H., Xiong Y. An experimental study on repair of peripheral nerve injury by transplantation of microcapsulated NGF-expressing NIH 3T3 cells // Zhonghua Zheng Xing Wai Ke Za Zhi. — 2005. — Vol. 1. — Р. 53-7.
21. Komobuchi H., Hato N., Teraoka M., Wakisaka H., Takahashi H., Gyo K., Tabata Y., Yamamoto M. Basic fibroblast growth factor combined with biodegradable hydrogel promotes healing of facial nerve after compression injury: an experimental study // Acta Otolaryngol. — 2010. — Vol. 130, № 1. — Р. 173-8.
22. Sondell M., Lundborg G., Kanje M. Vascular endothelial growth factor stimulates Schwann cell invasion and neovascularization of acellular nerve grafts // Brain Res. — 1999 Nov 6. — Vol. 846, № 2. — Р. 219-28.
23. Stanec S., Stanec Z. Reconstruction of upper-extremity peripheral-nerve injuries with ePTFE conduits // J Reconstr Microsurg. — 1998 May. — Vol. 14, № 4):227-32.
24. Lanzetta M., Gal A., Wright B., Owen E. Effect of FK506 and basic fibroblast growth factor on nerve regeneration using a polytetrafluoroethylene chamber for nerve repair // Int Surg. 2003. — Vol. 88, № 1. — Р. 47-51.
25. Borin A., Cruz O.L., Ho P.L., Testa J.R., Fukuda Y. The action of topical basic fibroblast growth factor in facial nerve regeneration // Otol Neurotol. — 2010 Apr. — Vol. 31, № 3. — Р. 498-505.
26. Toledo R.N., Borin A., Cruz O.L., Ho P.L., Testa J.R., Fukuda Y. The action of topical basic fibroblast growth factor in facial nerve regeneration // Otol Neurotol. 2010. — Vol. 31, № 3. — Р. 498-505.
27. Масгутов Р.Ф., Салафутдинов И.И., Богов А.А. (мл.), Трофимова А.А., Ханнанова И.Г., Муллин Р.И., Исламов Р.Р., Челышев Ю.А., Богов А.А., Ризванов А.А. Стимуляция посттравматической регенерации седалищного нерва крысы с помощью плазмиды, экспрессирующей сосудистый эндотелиальный фактор роста фибробластов. Клеточная трансплантология и тканевая регенерация. — 2011. — Т. VI, № 3.
28. Челышев Ю.А., Мухамедшина Я.О., Шаймарданова Г.Ф., Николаев С.И. Прямая доставка терапевтических генов для стимулирования посттравматической нейрорегенерации // Неврологический вестник. — 2012. — T. XLIV, вып. 1. — С. 76-83.
Многие психические расстройства, возникающие на органической почве , отличаются резистентностью к терапии, что , вероятно, обусловлено тем, что центральная нервная система — в отличие от периферической неспособна восстанавливать поврежденные нервные волокна или полноценно реагировать со структурной пластичностью. Нервные волокна нейронов головного мозга «пытаются» после его повреждения давать отростки , но им не удается восстанавливать их на большие расстояния и они не вступают в контакт с отростками других нейронов ( «клеток — мишеней»). Два фактора лежат в основе этой «неполноценности»: «внутреняя неспособность» центральных волокон к росту и негативное влияние факторов среды , окружающей эти волокна. Любой вариант повреждения аксонов вызывает множество серьезных молекулярных и клеточных ответов на уровне их тел и в месте повреждения. Повреждение нервных волокон может в некторых случаях привести к гибели тела нейрона. Для восстановления поврежденных аксонов — выживаемость тела нейрона и его способность вызывать экспрессию генов , что ведет к возобновлению роста и реиннервации первоначальных «клеток — мишений». Чем ближе к телу клетки аксональное повреждение , тем меньше у нейрона шансов выжить. Если аксоны повреждены вдали от тела нейрона , то последний хотя и уменьшается в размерах ( атрофия) , но все же выживает после такого повреждения, однако, им все равно не избежать значительной перестройки эндоплазматического ретикулума и аппарата Гольджи — хроматолиз, причем последний часто сопровождается увеличением экспрессии набора генов , связанного с ростом аксонов. Во всех случаях нейрон в конечном счете даже если и переживет повреждение , его нервные окончания и сегмент дистальный у очагу повреждения , будет дегенерировать ( вторичная дегенерация нервных волокон или Валлеровское перерождение).
Зародышевые аксоны центральной нервной системы восстанавливаются сравнительно легко , но со временем , с возрастом человека, они теряют такую способность. Взрослые нейроны утрачивают свою способность к росту уже во время послеродового развития , а их регенерационный ответ становится слабым и неполноценным. Утрата способности к росту совпадает со «взрослением» клеток нейроглии, астроцитов и олигодендроцитов и также с началом процесса миелинизации. Однако, миелин и связанные молекулы являются не единственными кандидатами , отвечающими за потерю регенерационной способности в процессе роста. В то же время, поврежденные нейроны посылают другим нейронам свои сигналы , вынуждая их терять свою способность к росту аксонов. Утрата регенерационной способности в процессе развития связывается и с внутренним уменьшением нейронной экспрессии белков , связанных с ростом, таких как GAP-43 и SCG10 и уменьшением уровней эндогенного циклического аденозинмонофосфата. К внешним факторам препятствующим росту аксонов относятся: клеточные компоненты ингибирования ( глиальный рубец, Валлеровское перерождение — олигодендроциты и миелин , вторичное повреждение : воспаление , клеточно — опосредованный иммунитет и потеря глиальных клеток; генетическая регуляция клеточных ответов на повреждение центральной нервной системы) , молекулярные компоненты ингибирования ( ингибиторы роста аксонов в глиальном рубце, хондроитинсульфатпротеингликаны, тенасцины, семафорины, миелин- ассоциированные ингибиторы роста, миелин — ассоциированный гликопротеин, олигодендроцит — миелиновый гликопротеин ) , передача сигналов , ингибирующих рост аксонов ( рецепторы для миелин — ассоциированных ингибиторов, сигнальные пути миелин — ассоиированных ингибиторов, циклический аденозинмонофосфат , протеинкиназа А). В последние годы достигнуты некоторые успехи в регенерации аксонов , в частности, с помощью введения нейротрофических факторов , блокаторов миелин — ассоциированных ингибиторов роста и / или их рецепторов.
Ряд психических растройств, в первую очередь, отличающихся нейродегенеративной природой , возникает вследствие неправильной компановки и накопления белка. Однако, сегодня еще не ясно : почему белок в этом случае приобретает определенную форму или претерпевает конформационные изменения и каким образом последние связаны с клиническими симптомами психического расстройства?
Категория сообщения в блог:
From Wikipedia, the free encyclopedia
Neuroregeneration refers to the regrowth or repair of nervous tissues, cells or cell products. Such mechanisms may include generation of new neurons, glia, axons, myelin, or synapses. Neuroregeneration differs between the peripheral nervous system (PNS) and the central nervous system (CNS) by the functional mechanisms involved, especially in the extent and speed of repair. When an axon is damaged, the distal segment undergoes Wallerian degeneration, losing its myelin sheath. The proximal segment can either die by apoptosis or undergo the chromatolytic reaction, which is an attempt at repair. In the CNS, synaptic stripping occurs as glial foot processes invade the dead synapse.[1]
Nervous system injuries affect over 90,000 people every year.[2] It is estimated that spinal cord injuries alone affect 10,000 each year.[3] As a result of this high incidence of neurological injuries, nerve regeneration and repair, a subfield of neural tissue engineering, is becoming a rapidly growing field dedicated to the discovery of new ways to recover nerve functionality after injury. The nervous system is divided into two parts: the central nervous system, which consists of the brain and spinal cord, and the peripheral nervous system, which consists of cranial and spinal nerves along with their associated ganglia. While the peripheral nervous system has an intrinsic ability for repair and regeneration, the central nervous system is, for the most part, incapable of self-repair and regeneration. There is currently no treatment for recovering human nerve function after injury to the central nervous system.[4] In addition, multiple attempts at nerve re-growth across the PNS-CNS transition have not been successful.[4] There is simply not enough knowledge about regeneration in the central nervous system. In addition, although the peripheral nervous system has the capability for regeneration, much research still needs to be done to optimize the environment for maximum regrowth potential. Neuroregeneration is important clinically, as it is part of the pathogenesis of many diseases, including multiple sclerosis.
Peripheral nervous system regeneration[edit]

Guillain–Barré syndrome – nerve damage
Neuroregeneration in the peripheral nervous system (PNS) occurs to a significant degree.[5][6] After an injury to the axon, peripheral neurons activate a variety of signaling pathways which turn on pro-growth genes, leading to reformation of a functional growth cone and regeneration. The growth of these axons is also governed by chemotactic factors secreted from Schwann cells. Injury to the peripheral nervous system immediately elicits the migration of phagocytes, Schwann cells, and macrophages to the lesion site in order to clear away debris such as damaged tissue which is inhibitory to regeneration. When a nerve axon is severed, the end still attached to the cell body is labeled the proximal segment, while the other end is called the distal segment. After injury, the proximal end swells and experiences some retrograde degeneration, but once the debris is cleared, it begins to sprout axons and the presence of growth cones can be detected. The proximal axons are able to regrow as long as the cell body is intact, and they have made contact with the Schwann cells in the endoneurium (also known as the endoneurial tube or channel). Human axon growth rates can reach 2 mm/day in small nerves and 5 mm/day in large nerves.[4] The distal segment, however, experiences Wallerian degeneration within hours of the injury; the axons and myelin degenerate, but the endoneurium remains. In the later stages of regeneration the remaining endoneurial tube directs axon growth back to the correct targets. During Wallerian degeneration, Schwann cells grow in ordered columns along the endoneurial tube, creating a band of Büngner cells that protects and preserves the endoneurial channel. Also, macrophages and Schwann cells release neurotrophic factors that enhance re-growth.
Central nervous system regeneration[edit]
Unlike peripheral nervous system injury, injury to the central nervous system is not followed by extensive regeneration. It is limited by the inhibitory influences of the glial and extracellular environment. The hostile, non-permissive growth environment is, in part, created by the migration of myelin-associated inhibitors, astrocytes, oligodendrocytes, oligodendrocyte precursors, and microglia. The environment within the CNS, especially following trauma, counteracts the repair of myelin and neurons. Growth factors are not expressed or re-expressed; for instance, the extracellular matrix is lacking laminins. Glial scars rapidly form, and the glia actually produce factors that inhibit remyelination and axon repair; for instance, NOGO and NI-35.[6][7][8] The axons themselves also lose the potential for growth with age, due to a decrease in GAP43 expression, among others.
Slower degeneration of the distal segment than that which occurs in the peripheral nervous system also contributes to the inhibitory environment because inhibitory myelin and axonal debris are not cleared away as quickly. All these factors contribute to the formation of what is known as a glial scar, which axons cannot grow across. The proximal segment attempts to regenerate after injury, but its growth is hindered by the environment. It is important to note that central nervous system axons have been proven to regrow in permissive environments; therefore, the primary problem to central nervous system axonal regeneration is crossing or eliminating the inhibitory lesion site.[4] Another problem is that the morphology and functional properties of central nervous system neurons are highly complex, for this reason a neuron functionally identical cannot be replaced by one of another type (Llinás’ law).[9]
Inhibition of axonal regrowth[edit]
Glial cell scar formation is induced following damage to the nervous system. In the central nervous system, this glial scar formation significantly inhibits nerve regeneration, which leads to a loss of function. Several families of molecules are released that promote and drive glial scar formation. For instance, transforming growth factors B-1 and -2, interleukins, and cytokines play a role in the initiation of scar formation. The accumulation of reactive astrocytes at the site of injury and the up regulation of molecules that are inhibitory for neurite outgrowth contribute to the failure of neuroregeneration.[10] The up-regulated molecules alter the composition of the extracellular matrix in a way that has been shown to inhibit neurite outgrowth extension. This scar formation involves several cell types and families of molecules.
Chondroitin sulfate proteoglycan[edit]
In response to scar-inducing factors, astrocytes up regulate the production of chondroitin sulfate proteoglycans. Astrocytes are a predominant type of glial cell in the central nervous system that provide many functions including damage mitigation, repair, and glial scar formation.[11] The RhoA pathway is involved. Chondroitin sulfate proteoglycans (CSPGs) have been shown to be up regulated in the central nervous system (CNS) following injury. Repeating disaccharides of glucuronic acid and galactosamine, glycosaminoglycans (CS-GAGs), are covalently coupled to the protein core CSPGs. CSPGs have been shown to inhibit regeneration in vitro and in vivo, but the role that the CSPG core protein vs. CS-GAGs had not been studied until recently.
Keratan sulfate proteoglycans[edit]
Like the chondroitin sulfate proteoglycans, keratan sulfate proteoglycan (KSPG) production is up regulated in reactive astrocytes as part of glial scar formation. KSPGs have also been shown to inhibit neurite outgrowth extension, limiting nerve regeneration. Keratan sulfate, also called keratosulfate, is formed from repeating disaccharide galactose units and N-acetylglucosamines. It is also 6-sulfated. This sulfation is crucial to the elongation of the keratan sulfate chain. A study was done using N-acetylglucosamine 6-O-sulfotransferase-1 deficient mice. The wild type mouse showed a significant up regulation of mRNA expressing N-acetylglucosamine 6-O-sulfotransferase-1 at the site of cortical injury. However, in the N-acetylglucosamine 6-O-sulfotransferase-1 deficient mice, the expression of keratan sulfate was significantly decreased when compared to the wild type mice. Similarly, glial scar formation was significantly reduced in the N-acetylglucosamine 6-O-sulfotransferase-1 mice, and as a result, nerve regeneration was less inhibited.[10]
Other inhibitory factors[edit]
Proteins of oligodendritic or glial debris origin that influence neuroregeneration:
- NOGO –The protein family Nogo, particularly Nogo-A, has been identified as an inhibitor of remyelination in the CNS, especially in autoimmune mediated demyelination, such as found in experimental autoimmune encephalomyelitis (EAE), and multiple sclerosis (MS). Nogo A functions via either its amino-Nogo terminus through an unknown receptor, or by its Nogo-66 terminus through NgR1, p75, TROY or LINGO1. Antagonising this inhibitor results in improved remyelination, as it is involved in the RhoA pathway.[6]
- NI-35 a non-permissive growth factor from myelin.
- MAG –Myelin-associated glycoprotein acts via the receptors NgR2, GT1b, NgR1, p75, TROY and LINGO1.
- OMgp –Oligodendrocyte myelin glycoprotein
- Ephrin B3 functions through the EphA4 receptor and inhibits remyelination.[6]
- Sema 4D(Semaphorin 4D) functions through the PlexinB1 receptor and inhibits remyelination.[6]
- Sema 3A (Semaphorin 3A) is present in the scar that forms in both central nervous system[12][13] and peripheral nerve injuries [14] and contributes to the outgrowth-inhibitory properties of these scars
Clinical treatments[edit]
Neurons replacement[edit]
in vivo glias to neurons reprogramming[edit]
Transcription factors, activation of genes (using CRISPR activation[15]) or small molecules are used to reprogram glias into neurons.
The most commonly targeted glias are astrocytes (usually using GFAP) because they share the same lineage as neurons and region—specific transcription signatures,[16] while the vector used is typically an adeno-associated virus because some serotypes pass the blood brain barrier and it does not cause disease.
Targeted genes usually depends on the type of neuron seeked (NGN2 is known to produce glutamatergic, ASCL1: GABAergic…) ; RBPJ-k block the Notch pathway and elicit a neurogenic program[17] and Sox2 can also increase reprograming efficiency by causing a dedifferentiation and self-amplification phase before maturating as neurons.
While theses techniques show lot of promises in animal models to many otherwise incurable neurodegenerative diseases and brain injuries, no clinical trials have started as of 2023.
Neural stem cells grafting[edit]
Tissue regrowth[edit]
Peripheral[edit]
Surgery[edit]
Surgery can be done in case a peripheral nerve has become cut or otherwise divided. This is called peripheral nerve reconstruction. The injured nerve is identified and exposed so that normal nerve tissue can be examined above and below the level of injury, usually with magnification, using either loupes or an operating microscope. If a large segment of nerve is harmed, as can happen in a crush or stretch injury, the nerve will need to be exposed over a larger area. Injured portions of the nerve are removed. The cut nerve endings are then carefully reapproximated using very small sutures. The nerve repair must be covered by healthy tissue, which can be as simple as closing the skin or it can require moving skin or muscle to provide healthy padded coverage over the nerve.[18] The type of anesthesia used depends on the complexity of the injury. A surgical tourniquet is almost always used.[18]
Prognosis[edit]
The expectations after surgical repair of a divided peripheral nerve depends on several factors:
- Age: Recovery of a nerve after surgical repair depends mainly on the age of the patient. Young children can recover close-to-normal nerve function. In contrast, a patient over 60 years old with a cut nerve in the hand would expect to recover only protective sensation; that is, the ability to distinguish hot/cold or sharp/dull.[18]
- The mechanism of injury: Sharp injuries, such as a knife wound, damage only a very short segment of the nerve, availing for direct suture. In contrast, nerves that are divided by stretch or crush may be damaged over long segments. These nerve injuries are more difficult to treat and generally have a poorer outcome. In addition, associated injuries, like injury to bone, muscle and skin, can make nerve recovery more difficult.[18]
- The level of injury: After a nerve is repaired, the regenerating nerve endings must grow all the way to their target. For example, a nerve injured at the wrist that normally provides sensation to the thumb must grow to the end of the thumb in order to provide sensation. The return of function decreases with increased distance over which a nerve must grow.[18]
Autologous nerve grafting[edit]
Currently, autologous nerve grafting, or a nerve autograft, is known as the gold standard for clinical treatments used to repair large lesion gaps in the peripheral nervous system. It is important that nerves are not repaired under tension,[18] which could otherwise happen if cut ends are reapproximated across a gap. Nerve segments are taken from another part of the body (the donor site) and inserted into the lesion to provide endoneurial tubes for axonal regeneration across the gap. However, this is not a perfect treatment; often the outcome is only limited function recovery. Also, partial de-innervation is frequently experienced at the donor site, and multiple surgeries are required to harvest the tissue and implant it.
When appropriate, a nearby donor may be used to supply innervation to lesioned nerves. Trauma to the donor can be minimized by utilizing a technique known as end-to-side repair. In this procedure, an epineurial window is created in the donor nerve and the proximal stump of the lesioned nerve is sutured over the window. Regenerating axons are redirected into the stump. Efficacy of this technique is partially dependent upon the degree of partial neurectomy performed on the donor, with increasing degrees of neurectomy giving rise to increasing axon regeneration within the lesioned nerve, but with the consequence of increasing deficit to the donor.[19]
Some evidence suggests that local delivery of soluble neurotrophic factors at the site of autologous nerve grafting may enhance axon regeneration within the graft and help expedite functional recovery of a paralyzed target.[20][21] Other evidence suggests that gene-therapy induced expression of neurotrophic factors within the target muscle itself can also help enhance axon regeneration.[22][23] Accelerating neuroregeneration and the reinnervation of a denervated target is critically important in order to reduce the possibility of permanent paralysis due to muscular atrophy.
Allografts and xenografts[edit]
Variations on the nerve autograft include the allograft and the xenograft. In allografts, the tissue for the graft is taken from another person, the donor, and implanted in the recipient. Xenografts involve taking donor tissue from another species. Allografts and xenografts have the same disadvantages as autografts, but in addition, tissue rejection from immune responses must also be taken into account. Often immunosuppression is required with these grafts. Disease transmission also becomes a factor when introducing tissue from another person or animal. Overall, allografts and xenografts do not match the quality of outcomes seen with autografts, but they are necessary when there is a lack of autologous nerve tissue.
Nerve guidance conduit[edit]
Because of the limited functionality received from autografts, the current gold standard for nerve regeneration and repair, recent neural tissue engineering research has focused on the development of bioartificial nerve guidance conduits in order to guide axonal regrowth. The creation of artificial nerve conduits is also known as entubulation because the nerve ends and intervening gap are enclosed within a tube composed of biological or synthetic materials.[24]
Immunisation[edit]
A direction of research is towards the use of drugs that target remyelinating inhibitor proteins, or other inhibitors. Possible strategies include vaccination against these proteins (active immunisation), or treatment with previously created antibodies (passive immunisation). These strategies appear promising on animal models with experimental autoimmune encephalomyelitis (EAE), a model of MS.[25]
Monoclonal antibodies have also been used against inhibitory factors such as NI-35 and NOGO.[26]
See also[edit]
- PTEN
- Muscle LIM protein
- Microtubule detyrosination
- Myelinogenesis
- Neuroprotection
- Spinal cord injury research
References[edit]
- ^ Kandel ER, Schwartz JH, Jessell TM (2003). «Chapter 55: The formation and regeneration of synapses». Principles of neural Science (fourth ed.). Cambridge: McGraw Hill. ISBN 978-0-8385-7701-1.
- ^ Stabenfeldt SE, García AJ, LaPlaca MC (June 2006). «Thermoreversible laminin-functionalized hydrogel for neural tissue engineering». Journal of Biomedical Materials Research Part A. 77 (4): 718–25. doi:10.1002/jbm.a.30638. PMID 16555267.
- ^ Prang P, Müller R, Eljaouhari A, Heckmann K, Kunz W, Weber T, Faber C, Vroemen M, Bogdahn U, Weidner N (July 2006). «The promotion of oriented axonal regrowth in the injured spinal cord by alginate-based anisotropic capillary hydrogels». Biomaterials. 27 (19): 3560–9. doi:10.1016/j.biomaterials.2006.01.053. PMID 16500703.
- ^ a b c d Recknor JB, Mallapragada SK (2006). «Nerve Regeneration: Tissue Engineering Strategies». In Bronzino JD (ed.). The biomedical engineering handbook (third ed.). Boca Raton, Fla.: CRC Taylor & Francis. ISBN 978-0-8493-2123-8.
- ^ Mahar M, Cavalli V (June 2018). «Intrinsic mechanisms of neuronal axon regeneration». Nature Reviews. Neuroscience. 19 (6): 323–337. doi:10.1038/s41583-018-0001-8. PMC 5987780. PMID 29666508.
- ^ a b c d e Yiu G, He Z (August 2006). «Glial inhibition of CNS axon regeneration». Nature Reviews. Neuroscience. 7 (8): 617–27. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
- ^ Bradbury EJ, McMahon SB (August 2006). «Spinal cord repair strategies: why do they work?». Nature Reviews. Neuroscience. 7 (8): 644–53. doi:10.1038/nrn1964. PMID 16858392. S2CID 11890502.
- ^ Bregman BS, Kunkel-Bagden E, Schnell L, Dai HN, Gao D, Schwab ME (November 1995). «Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors». Nature. 378 (6556): 498–501. Bibcode:1995Natur.378..498B. doi:10.1038/378498a0. PMID 7477407. S2CID 4352534.
- ^ Llinás RR (November 2014). «Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective». Frontiers in Cellular Neuroscience. 8: 320. doi:10.3389/fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ a b Zhang H, Uchimura K, Kadomatsu K (November 2006). «Brain keratan sulfate and glial scar formation». Annals of the New York Academy of Sciences. 1086 (1): 81–90. Bibcode:2006NYASA1086…81Z. doi:10.1196/annals.1377.014. PMID 17185507. S2CID 27885790.
- ^ Song I, Dityatev A (January 2018). «Crosstalk between glia, extracellular matrix and neurons». Brain Research Bulletin. 136: 101–108. doi:10.1016/j.brainresbull.2017.03.003. PMID 28284900. S2CID 3287589.
- ^ De Winter F, Oudega M, Lankhorst AJ, Hamers FP, Blits B, Ruitenberg MJ, Pasterkamp RJ, Gispen WH, Verhaagen J (May 2002). «Injury-induced class 3 semaphorin expression in the rat spinal cord». Experimental Neurology. 175 (1): 61–75. doi:10.1006/exnr.2002.7884. PMID 12009760. S2CID 39940363.
- ^ Mecollari V, Nieuwenhuis B, Verhaagen J (2014). «A perspective on the role of class III semaphorin signaling in central nervous system trauma». Frontiers in Cellular Neuroscience. 8: 328. doi:10.3389/fncel.2014.00328. PMC 4209881. PMID 25386118.
- ^ Tannemaat MR, Korecka J, Ehlert EM, Mason MR, van Duinen SG, Boer GJ, Malessy MJ, Verhaagen J (December 2007). «Human neuroma contains increased levels of semaphorin 3A, which surrounds nerve fibers and reduces neurite extension in vitro». The Journal of Neuroscience. 27 (52): 14260–4. doi:10.1523/JNEUROSCI.4571-07.2007. PMC 6673446. PMID 18160633.
- ^ Herrero-Navarro Á, Puche-Aroca L, Moreno-Juan V, Sempere-Ferràndez A, Espinosa A, Susín R, et al. (April 2021). «Astrocytes and neurons share region-specific transcriptional signatures that confer regional identity to neuronal reprogramming». Science Advances. 7 (15): eabe8978. doi:10.1126/sciadv.abe8978. PMC 8026135. PMID 33827819.
- ^ Herrero-Navarro Á, Puche-Aroca L, Moreno-Juan V, Sempere-Ferràndez A, Espinosa A, Susín R, et al. (April 2021). «Astrocytes and neurons share region-specific transcriptional signatures that confer regional identity to neuronal reprogramming». Science Advances. 7 (15): eabe8978. doi:10.1126/sciadv.abe8978. PMC 8026135. PMID 33827819.
- ^ Zamboni M, Llorens-Bobadilla E, Magnusson JP, Frisén J (October 2020). «A Widespread Neurogenic Potential of Neocortical Astrocytes Is Induced by Injury». Cell Stem Cell. 27 (4): 605–617.e5. doi:10.1016/j.stem.2020.07.006. PMID 32758425.
- ^ a b c d e f Payne SH (2001). «Nerve repair and grafting in the upper extremity». Journal of the Southern Orthopaedic Association. 10 (3): 173–189. PMID 12132829.
- ^ Kalantarian B, Rice DC, Tiangco DA, Terzis JK (October 1998). «Gains and losses of the XII-VII component of the «baby-sitter» procedure: a morphometric analysis». Journal of Reconstructive Microsurgery. 14 (7): 459–71. doi:10.1055/s-2007-1000208. PMID 9819092.
- ^ Tiangco DA, Papakonstantinou KC, Mullinax KA, Terzis JK (May 2001). «IGF-I and end-to-side nerve repair: a dose-response study». Journal of Reconstructive Microsurgery. 17 (4): 247–56. doi:10.1055/s-2001-14516. PMID 11396586.
- ^ Fansa H, Schneider W, Wolf G, Keilhoff G (July 2002). «Influence of insulin-like growth factor-I (IGF-I) on nerve autografts and tissue-engineered nerve grafts». Muscle & Nerve. 26 (1): 87–93. doi:10.1002/mus.10165. PMID 12115953. S2CID 38261013.
- ^ Shiotani A, O’Malley BW, Coleman ME, Alila HW, Flint PW (September 1998). «Reinnervation of motor endplates and increased muscle fiber size after human insulin-like growth factor I gene transfer into the paralyzed larynx». Human Gene Therapy. 9 (14): 2039–47. doi:10.1089/hum.1998.9.14-2039. PMID 9759931.
- ^ Flint PW, Shiotani A, O’Malley BW (March 1999). «IGF-1 gene transfer into denervated rat laryngeal muscle». Archives of Otolaryngology–Head & Neck Surgery. 125 (3): 274–9. doi:10.1001/archotol.125.3.274. PMID 10190798.
- ^ Phillips JB, Bunting SC, Hall SM, Brown RA (2005). «Neural tissue engineering: a self-organizing collagen guidance conduit». Tissue Engineering. 11 (9–10): 1611–7. doi:10.1089/ten.2005.11.1611. PMID 16259614.
- ^ Karnezis T, Mandemakers W, McQualter JL, Zheng B, Ho PP, Jordan KA, Murray BM, Barres B, Tessier-Lavigne M, Bernard CC (July 2004). «The neurite outgrowth inhibitor Nogo A is involved in autoimmune-mediated demyelination». Nature Neuroscience. 7 (7): 736–44. doi:10.1038/nn1261. PMID 15184901. S2CID 9613584.
- ^ Buffo A, Zagrebelsky M, Huber AB, Skerra A, Schwab ME, Strata P, Rossi F (March 2000). «Application of neutralizing antibodies against NI-35/250 myelin-associated neurite growth inhibitory proteins to the adult rat cerebellum induces sprouting of uninjured purkinje cell axons». The Journal of Neuroscience. 20 (6): 2275–86. doi:10.1523/JNEUROSCI.20-06-02275.2000. PMC 6772513. PMID 10704503.
Further reading[edit]
- Авторы
- Резюме
- Файлы
- Ключевые слова
- Литература
Тутуров А.О.
1
1 СамГМУ
Исследование современных технологий в восстановлении протяжённых дефектов нервов является важной фундаментальной задачей нейрохирургии и травматологии. Данные повреждения дифференцируются по сумме вовлеченных в патологический процесс окружающих тканей и компонентов нерва. Наиболее сложного, объёмного и, часто, безрезультатного лечения требуют именно травмы, приводящие к формированию больших диастазов, нервов. Причиной этому служит сочетание сразу нескольких ключевых деталей: полное пересечение нервного ствола, потеря направляющего канала для регенерации, и, в дальнейшем, создание условий для образования посттравматической невромы. Тотальный разрыв всей полости, иначе говоря, невротмезис, приводит к утрате сообщения между нервными волокнами проксимального и дистального участка нерва, нарушает целостность миелиновой оболочки и соединительнотканных элементов. Вследствие разобщения волокон , потери направляющего канала – эпиневрия и утраты его внутренней среды регенерация не наступает. Все эти процессы приводят к образованию соединительнотканных колбовидных утолщений – невром, что указывает на терминальную стадию повреждения нерва – прекращения его функционирования в дистальном отделе. Исходя из анатомического обоснования сформированного диастаза ушивание дефекта эпиневральными швами не возможно, так как это неминуемо приведет к его перенатяжению или расслоению. Начиная с 1910-ых годов, многие нейрохирурги поставили перед собой задачу лечения данных травм. Развитие технологий направило исследования на создание кондуитов — искусственных проводников, помещаемых в диастаз нерва и создающих необходимые условия для регенерации нервных волокон в заданном направлении, за счет искусственной внутренней среды. С тех пор восстановление протяженных дефектов нервов стало возможным и получило широкое применение и развитие по всему миру. В обзоре проанализированы различные варианты кондуитов с их уникальной внутренней средой, а также смежные с ними разработки, показывающие использование регенераторного потенциала нервной ткани, для выявления наиболее верного направления в развитии технологий восстановления протяжённых дефектов нервов.
невротмезис
повреждение нерва
протяженные дефекты
кондуиты
регенераторный потенциал.
1. Берснев В.П. Хирургия позвоночника, спинного мозга и периферических нервов / Е.А. Давыдов, Е.Н. Кондаков // Специальная литература. — СПб, 1998. — С. 368.
2. Кубицкий А.А. Хирургическое лечение повреждений периферических нервов верхней конечности методами тракционного удлинения и аутонервной пластики: автореф. дис. … канд. мед. наук / А.А. Кубицкий.— Казань, 2002. — 24 с.
3. Козлов А. В., Афанасьев Л. М., Якушин О. А., Молочков Е. В. Хирургическая тактика лечения больных с повреждениями нервов // III съезд нейрохирургов России : материалы съезда. – СПб., 2002. – С. 526–527.
4. Бабаханов, Ф.Х. Микрохирургические реконструктивно-восстановительные операции при массивных повреждениях периферических нервов / Ф.Х.Бабаханов, А.К.Муминов, Д.Б.Якубов, Б.А.Абдушукуров // Материалы III съезда нейрохирургов России. М., 2002. — С. 52.
5. Патент РФ № 2153295. Способ аутонейропластики.
6. Патент РФ № 2179828. Способ выделения и отсечения икроножного нерва с образующими его ветвями и инструмент для его осуществления
7. Патент РФ №2154431. Способ лечения обширных повреждений лучевого нерва на плече.
8. Патент РФ №99107088/14. Способ восстановления чувствительности на стопе.
9. Борода Ю.И. Хирургия дефектов нервных стволов конечностей / Ю.И. Борода, В.П. Берснев // Мат. III съезда нейрохирургов России СПб, 2002. — С. 524525.
10.BY №10011. Способ восстановления нервного ствола.
11.Патент РФ №2471435. Способ восстановления поврежденного нерва при дефектах на большом протяжении.
12. Патент РФ №2176482. Способ пластики дефекта нервного ствола.
13. Senoglu M., Nacitarhan V., Kurutas E.B. Intraperitoneal Alpha-Lipoic Acid to prevent neural damage after crush injury to the rat sciatic nerve / M. Senoglu [etc.] // J Brachial Plex Peripher Nerve Inj. — 2009. — P. 22.
14. Патент РФ №2347539. Способ лечения повреждения периферического нерва.
15. SU №1204197. Способ восстановления нервного ствола.
16. Foramitti C. Zur technik der nervennaht / C. Foramitti [etc.] // Arch Klin Chir. — 1904. — P. 643–648.
17. Tang J.B., Gu Y.Q., Song Y.S. Repair of digital nerve defect with autogenous vein graft during flexor tendon surgery in zone 2 / J. B. Tang [etc.] // J Hand Surg Br. — 1993. — P. 449-453.
18. Chiu D.T., Strauch B. A prospective clinical evaluation of autogenous vein grafts used as a nerve conduit for distal sensory nerve defects of 3 cm or less / D.T. Chiu [etc.] // Plast Reconstr Surg. — 1990. — P. 928-934.
19. Walton R.L., Brown R.E., Matory W.E. Autogenous vein graft repair of digital nerve defects in the finger: a retrospective clinical study / R.L. Walton[etc.] // Plast Reconstr Surg. – 1989. – P. 944-949.
20.Pateman C. J., Harding A. J., Glen A. Nerve guides manufactured from photocurable polymers to aid peripheral nerve repair / C. J. Pateman [etc.] // Biomaterials. – 2015. — Vol. 49. – Р. 77-89.
21.Yamanaka T., Hosoi H., Murai T. Regeneration of the Nerves in the Aerial Cavity with an Artificial Nerve Conduit -Reconstruction of Chorda Tympani Nerve Gaps / T. Yamanaka [etc.] // PLOS ONE. – 2014. – Vol. 9, №4. — P. 1-7.
22. Rotshenker S. Wallerian degeneration: the innate-immune response to traumatic nerve injury / S. Rotshenker [etc.] // J Neuroinflammation. — 2011. — P. 109.
23. Wilson A.D., Hart A., Brännström T. Delayed acetyl-L-carnitine administration and its effect on sensory neuronal rescue after peripheral nerve injury / A.D. Wilson [etc.] // J Plast Reconstr Aesthet Surg. — 2007. – P. 114-118.
24. Dokkaew J. Vitamin E reduces sensory neuronal loss and improves nerve regeneration after sciatic nerve injury / J. Dokkaew [etc.] // Asian Biomed. — 2013. — P. 649-655.
25. Azizi S., Heshmatian B., Amini K. Alpha-lipoic acid loaded in chitosan conduit enhances sciatic nerve regeneration in rat / S. Azizi [etc.] // Iran J Basic Med Sci. — 2015. — Vol. 18, №3. –– Р. 229-234.
26. Leighton J., Mansukhani S., Estes L. Decalcifiied eggshell membrane, a supporting substrate for electron microscopic cross sections of monolayers of epithelial cell line MDCK / J. Leighton [etc.] // In Vitro. – 1971 – P. 251-252.
27. Ruff K.J., Winkler A., Jackson R.W. Eggshell membrane in the treatment of pain and stiffness from osteoarthritis of the knee: a randomized, multicenter, double-blind, placebo-controlled clinical study / K.J. Ruff [etc.] // Clin Rheumatol. – 2009. – P. 907-914.
28. Cordeiro C.M., Hincke M.T. Recent patents on eggshell: shell and membrane applications / C.M. Cordeiro [etc.] // Recent Pat Food Nutr Agric. – 2011. – P. 1-8.
29. Jia J., Duan Y.Y., Yu J. Preparation and immobilization of soluble eggshell membrane protein on the electrospun nanofibers to enhance cell adhesion and growth // J. Jia [etc.] // J Biomed Mater Res A. – 2008. – P. 346-373.
30. Arias J.I., Gonzalez A., Fernandez Ms. Eggshell membrane as a biodegradable bone regeneration inhibitor / J.I. Arias [etc.] // J Tissue Regen Med. – 2008, — P. 228-235.
31.Farjah Gh, Heshmatian B, Karimipour M, Saberi A. Using Eggshell Membrane as Nerve Guide Channels in Peripheral Nerve Regeneration / Gh.Farjah [etc.] // Iran J Basic Med. — 2013. Sci. Vol.16, №8. – P. 901-905.
32. United States Patent №20120150205 A1. Silk nanofiber nerve conduit and method for producing thereof.
33. United States Patent №20050013844 A1. Neural regeneration conduit.
34. Zhang J., Sheng L., Jin C. Liquid Metal as Connecting or Functional Recovery Channel for the Transected Sciatic Nerve / J. Zhang [etc.] // Beijing, China. – 2014.
http://medicalxpress.com/news/2014-04-beijing-explore-liquid-metal-reconnect.html
35. Патент РФ №2471435. Способ восстановления поврежденного нерва при дефектах на большом протяжении.
36. United States Patent №20120109167 A1. Nerve guide conduit containing carbon nanotubes.
37. United States Patent №20030176876 A1. Multi-channel bioresorbable nerve regeneration conduit and process for preparing the same.
1. Введение
Проблема лечения больных с повреждениями периферических нервов является одной из ведущих в травматологии и нейрохирургии. Нарушения целостности нервов сопровождают 3-10% от всех травм опорно-двигательного аппарата и приводят к стойкой утрате трудоспособности у 60-63% пациентов, более того, почти 80% этой группы составляют лица молодого трудоспособного возраста [1, 2]. В большинстве случаев травма нервов, вследствие специфики анатомо-топографических взаимоотношений, редко бывает изолированной [3].
Рубцовые изменения, а так же дополнительные повреждения нервов при неквалифицированном хирургическом вмешательстве, ущемления между костными отломками и другие обстоятельства приводят к возникновению большого диастаза между фрагментами поврежденного нерва. К их числу чаще относятся травмы в диафизарных отделах конечностей, где различные методы сгибания соответствующих суставов с целью уменьшения натяжения нерва при его сшивании не дают результатов [4].
Современные методы восстановления протяженных дефектов нервов, можно разделить на три большие группы, дифференцированные по виду хирургического вмешательства и особенностей его технологии:
1- Аутотрансплантация нерва;
2- Аутотрансплантация тканей для создания направления регенерации нерва;
3- Создание искусственных проводников и (или) ксенотрансплантатов для создания направления регенерации нерва
Выбор оптимального вида реконструктивного вмешательства по восстановлению непрерывности нервного ствола должен базироваться не только на величине дефекта нерва, но также учитывать совокупность факторов, среди которых локализация повреждения, способ сближения его концов и объективная оценка степени натяжения в зоне шва.
2. Аутотрансплантация нервов и тканей для создания направления регенерации нерва.
В связи с решением перечисленных вопросов разработаны многие методики, являющиеся смежными по отношению к основной теме обзора. Например, способ сшивания концов нерва, позволяющий уменьшить натяжение в зоне шва, с помощью эпиневральных лоскутов. Данный способ аутопластики дефекта нервного ствола заключается в расположении трансплантатов между концами нерва с учетом их анатомического строения, сближении и фиксации концов нерва за эпиневральные лоскуты к подлежащим тканям, а также обкладывании трансплантатов фрагментами подкожно-жировой клетчатки на сосудистой ножке [5]. Трансплантаты готовятся с помощью специальных устройств, позволяющих выделять и отсекать участки икроножного нерва с образующими его ветвями [6]. Широко применяется метод лечения повреждений нервов посредством восстановления их функций за счёт подшивания повреждённого нерва через аутотрансплантат к нормально функционирующим нервам той же части конечности [7-9].
Довольно интересной считается методика[10], по мнению авторов которой сшивание концов нерва с наложением поперечных насечек эпиневрия и периневрия в шахматном порядке или циркулярно, позволяет обеспечить зону восстановления нерва необходимыми трофическими ресурсами.
Известен способ пластики дефекта нервного ствола [11], который заключается в том, что в сосудисто-нервный комплекс с манжетой из выкроенного из подлежащей собственной фасции голени фасциально-клетчаточного островкового лоскута включают икроножный нерв с малой подкожной веной в одном блоке. Таким образом, за счет перемещения целой системы тканей в посттравматический диастаз, можно говорить о создании сложного кондуита. Данную разработку практически повторяет метод [12], в котором, как проводник используется выделенный трансплантат в виде сосудисто-нервного комплекса из икроножного нерва с малой подкожной веной в одном блоке, перенесенный в дефект нерва.
Но, не смотря на вышеперечисленные работы, при травме с дефектом длины нервных стволов, а также при реконструкции по поводу застарелых повреждений, для сближения концов требуется: 1) значительное его натяжение; 2) сгибание в смежных суставах; и как альтернатива — 3) аутонервная пластика. Частыми осложнениями аутопластики являются: натяжение нерва, приводящее к «прорезыванию швов»; сгибание в смежных суставах, вследствие которого развиваются контрактуры в послеоперационном периоде и вторичное повреждение нервов при разработке движений; ротация нервных волокон в различных плоскостях, что препятствует их обособлению в пучки. Более того, при аутонервной пластике приходится использовать нерв с другого участка тела, что приводит к излишней анестезии в зоне иннервации, формированию дополнительной полости с возможными осложнениями, а также затрате времени на хирургическое вмешательство. Все это значительно ухудшает результаты восстановления функции нервного ствола, причем регенераторный потенциал в вышеперечисленных методиках используется не полностью.[13]
3. Создание искусственных проводников и (или) ксенотрансплантатов для создания направления регенерации нерва
Одной из исследовательских работ, оказавших влияние на развитие создания искусственных проводников, на наш взгляд, может стать способ лечения повреждения периферического нерва [14], который заключается в формировании изолирующего кондуита, выполненного из тонкостенной силиконовой трубки. Длину силиконовой трубки выбирают такой, чтобы она была на 4-5 см больше размера восстановленного дефекта нервного ствола. После восстановления целостности поврежденного нерва, а именно ушивания дефекта, изолирующую муфту продвигают проксимально до перекрытия изолирующей муфтой на 4-5 см места соединения проксимального и дистального концов нерва. В данном исследовании нет указания на использование силикона, как кондуита, помещенного в диастаз посттравматического нерва для направления регенерации, но идея использования трубки из инородной ткани, как изолятор для регенерации сшитого нерва, может считаться первой ступенью на пути к дальнейшему развитию нового направления в восстановительной медицине.
Совершенно новым методом восстановления протяженных дефектов нервных стволов является замещение диастаза нерва проводником, содержащим внутреннюю среду, повторяющую естественные условия регенерации.
Одним из прорывных трудов в развитии данного направления стал “Способ восстановления нервного ствола” [15], который заключается в соединении проксимального и дистального концов повреждённого нерва с трансплантированным сосудом. С целью ускорения восстановления целостности и достижения полноты регенерации, в качестве трансплантата авторы использовали артериальный сосуд донора, заполненный гомогенатом леммоцитов. В просвет трансплантата с двух сторон вводили концы нервного ствола и соединяли биоклеем их эпиневральные оболочки с краями трансплантата. Клетки Шванна обеспечивают направленных рост регенерирующих аксонов перерезанного нерва от проксимального конца к дистальному. Они образуют, так называемые “мостики регенерации”, по которым и происходит продвижение регенераторного процесса.
В современной врачебной практике в качестве транплантата между отрезками нерва более широкое применение получили – венозные проводники [16]. Удовлетворительные функциональные результаты были получены в случае лечения дистальных чувствительных нервных дефектов до 3 мм. Выявлены несколько преимуществ использования вены, как проводника между концами пересечённого нерва (Chiu etc. 1982): базальная мембрана, которая образует эндотелий вены, богата ламинином, а адвентиция состоит в основном из коллагена; стенки вен являются достаточно упругими, чтобы действовать в качестве барьера против рубцового разрастания и имеют проницаемость, чтобы обеспечить диффузию питательных веществ внутрь (Wang etc., 1993; Tseng etc. 2003). Сторонники данного вида проводников отмечают достаточный запас для выбора венозных шунтов из многих областей тела, низкой стоимости и относительно малой степенью дополнительного повреждения в сравнении с нервными трансплантатами. Кроме того, вены неиммуногенны, практически не вызывают воспалительную реакцию, и доступны в широком разнообразии размеров. Они могут обеспечить механическую поддержку регенерирующих аксонов, предлагая защищенную биохимическую среду, вдали от окружающих тканей (Allet др., 2003).
Но венозные кондуиты имеют и существенные недостатки. В течение процесса регенерации, они оказались нестойкими, что приводило к перегибам или распадам стенки трубки [17-19]. По этой причине их использование становится невозможным при восстановлении больших промежутков диастаза нерва. Чиу с соавторами были проведены исследования, доказавшие тот факт, что вена, как проводник, может использоваться лишь при немногих травмах нервов, в частности, при диастазах длиной менее 3 мм [18].
В настоящее время развивается совершенно новое направление в восстановлении протяженных дефектов нервов, а именно создание искусственных проводников. Это направление исследований считается более перспективным, чем аутотрансплантация, поскольку их результатом станет разработка метода восстановления протяжённых повреждений нервного ствола. Исследования направлены на разработку новых возможностей при конструировании кондуита, в которые входят подборка индивидуального диаметра и анатомической формы под стать поврежденному нерву, использование биоразлагаемых материалов, применение всякого рода направляющих борозд в его стенке для более точного сопоставления нервных волокон и контроля регенераторных процессов.
Ученые Университета Шеффилда разработали и испытали метод изготовления проводника с помощью микростереолитографии. Это технология 3D-печати, позволяющая точно воссоздать микроструктуру нерва. Испытание изготовленного устройства было проведено на мышах со значительной травмой нерва: животному восстановили нервное волокно длиной около 3 мм за 21 день. Для изготовления устройства был использован полиэтиленгликоль, который одобрен для клинического использования. Материал не является биоразлагаемым, что создаёт некоторые трудности при применении данного типа кондуита. Стоит упомянуть о двух важных особенностях, а именно: наличие на внутренней поверхности стенки проводника особых насечек, которые способствуют более точному направлению нервных волокон в трубке, и свойстве данного материал проводить нервный импульс, что даёт право отнести его к биоинертным[20].
Итак, как отмечено выше, для создания направляющих кондуитов лучше всего подойдут биоразлагаемые материалы. Наиболее перспективным представляется использование коллагена, полигликолевой кислоты (ПГК) и их конгломерата.
Очень интересным не только с точки зрения конструкции кондуита, но и его содержимого является сочетание данных материалов. База трубки была изготовлена из цилиндрически тканой ПГК сетки, внешняя и внутренняя поверхности которой были покрыты аморфным слоем коллагена. Просвет трубки был наполнен 3D-губчатой коллагеновой матрицей. Губчатый коллаген обеспечивает благоприятную микросреду для восстановления нерва (регенерации аксонов, клеточной пролиферации и заживления тканей). Несмотря на то, что коллаген имеет тенденцию растворяться в организме, ПГК-кондуит позволяет сохранять его форму в течение достаточно долгого времени для полной регенерации нерва.
ПГК-коллаген трубка была 30 мм в длину и 0,7 мм в диаметре, что позволило перекрыть дефект нерва с небольшим запасом для наложения эпиневральных швов нерв-кондуит.[21]
Как было сказано выше, коллаген является хорошим материалом не только для создания направляющей трубки при повреждениях нерва, но и искусственных условий регенерации нервной ткани, т.е. внутренней среды для кондуита. Кроме него целесообразным считается использование альфа-липоевой кислоты(АЛК).
Известно, что повреждение нерва связано с окислительным стрессом. Увеличение уровня свободных радикалов кислорода и снижение деятельности ферментов антиоксидантной защиты наблюдаются после травмы нерва. Антиоксиданты, такие как ацетил-L-карнитин, витамин Е, АЛК успешно используются в лечении экспериментального пересечения нерва. Однако, эффект АЛК в регенерации периферических нервов до конца не выяснены.[22-24]
Совсем недавно исследовали два совершенно новых материала для создания кондуитов. Это хитозан и мембрана яичной скорлупы.
Хитозан – это аминосахар, производное линейного полисахарида, одним из источников получения которого служат панцири ракообразных. Успешным применением данного материала служит исследование с заполнением просвета хитозанового проводника альфа-липоевой кислотой (обоснование её использования указаны выше)[25].
Мембрана яичной скорлупы оказалась невероятно подходящим материалом для формирования трубки. Путём многократной обработки яичной скорлупы: сделав отверстие в скорлупе, вынуть содержимое; промыть изнутри, погрузить в 5% уксусную кислоту на 8 дней, далее в фосфатно-солевой буфер ph 7.4 на 30 минут; надеть на тефлоновую трубку; высушить 37 градусов 24 часа в стерильном ламинарном боксе; [26] была получена тонкая пленка. Данная мембрана сохраняет белок и предотвращает проникновение бактерий. Мембрана яичной скорлупы является естественным источником комбинированных глюкозамина, хондроитина и гиалуроновой кислоты, [27] имеют высокое содержание биологически активных компонентов, а также способности к удержанию влаги, которые имеют потенциальную пользу.[28] Мембрана яичной скорлупы является нетоксичной [29] и биологически разлагаемой [30]. Это может иметь большой потенциал для восстановления нервов в исследованиях регенерации аксонов в периферической нервной системе [28].
Положительным результатом использования мембраны послужили следующие факты: отсутствие послеоперационных смертей, клинических признаков раневой инфекции. Операции были хорошо перенесены животными. Нет клинических признаков инфекции, болей или дискомфорта во время регенеративного периода. [31]
3.1. Кондуиты для восстановления повреждений многопучковых нервов
Развитие создания проводников из искусственных материалов привело исследователей к мысли о восстановления многопучковых нервов, например, седалищного. Идея заключается в помещении не одного кондуита, общего для всех нервных пучков, в диастаз нерва, а именно, нескольких пучковых проводников, заключенных в одну общую муфту. Также, как оказалось, вышеперечисленные материалы не являются финальной точкой в исследованиях, например, появился способ восстановления нервного ствола с помощью кондуита из шелковых нановолокон. Кондуит создавали путем электроформования волокнистого прядильного раствора в цилиндре, смазанного изнутри полиэтиленоксидом. Электроформование — это процесс создания волокон, который отличается определенной гибкостью, позволяющей задавать конечные характеристики и структуру нановолокнам под стать параметрам нервного ствола.[32]
Говоря о параметрах и структуре изготавливаемых кондуитов, нельзя не упомянуть о способе восстановления нерва с помощью кондуита, имеющего спиралевидную геометрию.[33] Регенерация при выполнении данной методики осуществляется благодаря наличию различных клеток и биовеществ на каждом из слоёв скрученного из плоского листа кондуита, что также направляет разобщенные нервные волокна.
Немного другим подходом к решению поставленной задачи воспользовались авторы способа восстановления нерва [34] с помощью установки кондуита из жидкого сплава селена, галия и индия с формированием микроканальной структуры для направленного продвижения нервных волокон. К сожалению, наличие жидкого металла в структуре нерва отрицательно сказывается на его физиологии. Учтя это, выдвинули свои методы и другие группы ученых. Итогом исследований стало создание способа восстановления поврежденного нерва при дефектах на большом протяжении.[35] Восстановление происходит путём имплантации электропроводящих элементов из металла или полимерного токопроводящего материала, которые вводят в пучок нерва на 3-4 мм в оба конца диастаза нерва, далее весь диастаз укрывают кондуитом. Данная разработка чем-то напоминает взгляды на регенерацию нерва китайских исследователей, но использование кондуита выводит разработку российских авторов на качественно новый уровень.
На наш взгляд, способ восстановления нервного волокна [36], который заключается в создании проводника, имеющего две поверхности: внешнюю – силиконовую, и внутреннюю – из углеродных нанотрубок, мог бы стать вершиной восстановления нервов посредством кондуитов, но способ восстановления нервного ствола [37], который заключается в создании кондуита из пористого многоканального биоразлагаемого кондуита, помещаемого вместо диастаза нерва, внес свою весомую лепту в историю развития проводников между проксимальными и дистальными концами поврежденных нервов. По нашему мнению, данная форма кондуита универсальна и наиболее физиологична для восстановления нервного ствола. Остается только подобрать нужную ткань для его создания.
Безусловно, направление каждого пучка отдельно от других является наиболее верным методом восстановления повреждённого нерва и, определенно, через некоторое время он вытеснит все вышеперечисленные варианты использования регенераторного потенциала нерва.
Заключение
Итого, регенераторный потенциал нервного волокна можно использовать в искусственной среде, направляя и стимулируя его процессы.
В дальнейшем, технологию восстановления протяженных дефектов нервов с помощью биоразлагаемого кондуита можно направить и на восстановление спинного мозга, а, возможно, и — головного. К примеру, одно из самых частых осложнений при операции по удалению менингиомы — доброкачественной опухоли головного мозга, является повреждение обонятельного нерва, проходящего вблизи опухоли. При успехе данного метода, можно свести на нет один из рисков данной операции. И это лишь один из многих примеров прямого участия данного способа восстановления нервов.
В заключении хотелось бы сказать, что данная методика восстановления протяженных дефектов нервных волокон является прорывом в современной медицине и может найти широкое применение не только в нейрохирургии, но и травматологии, трансплантологии и других отраслях медицины
Библиографическая ссылка
Тутуров А.О. СОВРЕМЕННЫЕ ТЕНДЕНЦИИ В ВОССТАНОВЛЕНИИ ПРОТЯЖЁННЫХ ДЕФЕКТОВ НЕРВОВ. РАЗВИТИЕ ТЕХНОЛОГИЙ ИСПОЛЬЗОВАНИЯ РЕГЕНЕРАТОРНОГО ПОТЕНЦИАЛА НЕРВНЫХ ВОЛОКОН, ИХ СУЩНОСТЬ, НЕДОСТАТКИ И ПРЕИМУЩЕСТВА. // Международный студенческий научный вестник. – 2017. – № 1.
;
URL: https://eduherald.ru/ru/article/view?id=16825 (дата обращения: 05.04.2023).
Предлагаем вашему вниманию журналы, издающиеся в издательстве «Академия Естествознания»
(Высокий импакт-фактор РИНЦ, тематика журналов охватывает все научные направления)
Одновременно с процессом денервации, возникшей при повреждении нерва, начинаются восстановительные процессы, которые могут идти по трем направлениям.
(1) Регенерация нерва: проксимальная культя формирует аксональные выросты (наплывы аксоплазмы, или «колбы роста»), которые начинают продвигаться дистально и врастать в эндоневральные трубки (разумеется, лишь в тех случаях, когда последние сохранили свою целостность). Миелиновая оболочка вновь образующегося волокна формируется из тяжей леммоцитов. Скорость регенерации аксонов составляет приблизительно 1.5–2 мм в день. Отдельные нервные проводники имеют различную способность к регенерации: среди периферических нервов особенно хорошо восстанавливается функция лучевого и мышечно-кожного нервов, а наихудшими регенеративными способностями обладают локтевой и малоберцовый нервы. Для достижения хорошего восстановления растущие аксоны должны подключиться к дистальной культе нерва до того, как в ней наступят выраженные пери- и эндоневральные рубцовые сращения. В случаях формирования по ходу прорастающего волокна соединительнотканного рубца часть аксонов не распространяется в дистальном направлении, а беспорядочно отклоняется в стороны, формируя травматическую неврому. При полном анатомическом повреждении нервного ствола через 2–3 недели после травмы на центральном конце образуется ампутационная неврома. Регенерация нервного ствола может происходить гетерогенно: часть двигательных волокон врастает в оболочки чувствительных, а одноименные — в пучки, иннервирующие разноименные участки конечности.
(2) В случаях, когда в нервном стволе поражаются не все, а лишь часть нервных волокон, восстановление функции мышц возможно за счет разветвления сохранившихся аксонов и «захвата» ими тех мышечных волокон, которые иннервировались погибшими аксонами; при этом происходит укрупнение двигательных единиц мышцы. За счет этого механизма мышца может сохранять свою работоспособность в случаях утраты до 50 % иннервировавших ее аксонов (а для мышц, не развивающих значительных усилий — даже до 90 %), однако для полного завершения процесса компенсаторной иннервационной перестройки требуется около года.
(3) В ряде случаев (обычно при травме нервного ствола типа ушиба) восстановление или улучшение функций связано с обратимостью некоторых патоморфологических процессов: с исчезновением реактивных воспалительных явлений, с рассасыванием мелких кровоизлияний и т. д. При нетяжелых травмах проводимость нерва, даже после полной ее потери, восстанавливается в течение первых дней или недель.
К основным факторам, определяющим при периферических невропатиях и плексопатиях скорость и степень спонтанного восстановления нарушенных функций (и, следовательно, объем и направленность лечебных вмешательств), относятся следующие:
‒ степень повреждения нервного проводника;
‒ уровень поражения;
‒ характер повреждающего агента.
Реабилитологи чаще всего определяют степень повреждения нерва по 3 категориям согласно классификации H.Seddon. Иногда используют также классификацию S.Sunderland, выделяющего 5 степеней повреждения нервов; эта классификация основана на классификации H.Seddon, детализируя ее. Согласно классификации H.Seddon, все локальные повреждения нервных стволов делят, в зависимости от сохранности аксона и соединительнотканных структур, на три группы:
‒ нейрапраксия;
‒ аксонотмезис;
‒ нейротмезис.
Нейрапраксия (neurapraxia, англ.) — это повреждение нерва, не приводящее к гибели аксона. Часто наблюдается при компрессии нерва (например, «ночной субботний паралич» вследствие компрессии лучевого нерва), при легкой травме нерва. Клинически характеризуется снижением вибрационной, проприоцептивной, иногда тактильной чувствительности. Болевая чувствительность страдает реже. Часто наблюдаются двигательные нарушения и парестезии. Блок проведения нервного импульса, наблюдающийся вследствие локального повреждения миелиновой оболочки, носит преходящий характер и регрессирует по мере восстановления миелина. Восстановление двигательных и чувствительных функций может продолжаться до 6 месяцев.
Аксонотмезис (axonotmesis, англ.) — повреждение нерва, приводящее к гибели аксона при сохранности эпиневрия, периневрия, эндоневрия и шванновских клеток. Нередко наблюдается при закрытых переломах либо вывихах костей конечностей, а также при сдавлениях нервных стволов. Нарушаются двигательные, чувствительные и судомоторные функции нерва. Восстановление функций происходит за счет регенерации аксона. Скорость и степень восстановления зависит от уровня поражения, возраста (у молодых регенерация происходит быстрее) и общего состояния больного. В случаях, когда прорастание аксона происходит медленно, может произойти рубцевание эндоневральной трубки, в которую прорастает аксон, и восстановление не наступает. По этой же причине неблагоприятный прогноз имеется в случаях, когда дефект нервного ствола имеет значительную длину. При благоприятных условиях происходит постепенная невротизация дистального отдела поврежденного нерва, которая продолжается в течение многих месяцев, иногда год и более. Наблюдается восстановление утраченных функций, но не всегда полное.
Нейротмезис (neurotmesis, англ.) — разрыв нерва с пересечением аксона и соединительнотканных оболочек нерва. Из-за того, что повреждаются эндоневральные трубки, невозможным становится прорастание в них аксонов, регенерация аксонов приводит к образованию травматической невромы. Прогноз восстановления неблагоприятный.
Данная классификация основана на микроскопических изменениях в нервном стволе. Макроскопически различить степень повреждения практически невозможно. Диагностика основана на динамическом клиническом и электрофизиологическом наблюдении. В связи с этим при закрытых травмах нервных стволов нередко применяют иную классификацию, основанную на выделении следующих 4 форм поражения нервного ствола:
‒ сотрясение
‒ ушиб
‒ сдавление
‒ тракция
Сотрясение не сопровождается морфологическими изменениями в нерве, нарушения функции нерва кратковременны (не более 1–2 недель) и полностью обратимы.
Ушиб нерва характеризуется возникновением мелких кровоизлияний, участков размозжения нервных волокон и пучков, что приводит к полному либо частичному нарушению проводимости, длительным и стойким выпадением функций.
При сдавлении нерва степень нарушения проводимости зависит в первую очередь от длительности сдавления: при своевременном удалении сдавливающих нерв субстратов (гематома, инородное тело, отломок кости и т. д.) может наблюдаться быстрое и полное восстановление проводимости, тогда как при длительном сдавлении в нервном стволе развиваются дегенеративные изменения. Отсутствие восстановления функции в течение 2–3 месяцев является критерием полного анатомического перерыва нерва.
Тракция (например, тракдия ветвей плечевого сплетения при вправлении вывиха плеча) обычно сопровождается частичным нарушением функции, однако восстановление проводимости по нерву происходит достаточно длительно (в течение нескольких месяцев).
Чем проксимальнее поражение нервного ствола или сплетения (т. е. чем больше расстояние от места повреждения до периферических окончаний), тем хуже прогноз восстановления функции, поскольку тем больший срок требуется для прорастания нервного волокна и тем больше вероятность развития в эндоневральной трубке периферического отрезка нерва необратимых рубцовых изменений. Так, например, по данным С. И. Карчикяна, при ранениях седалищного нерва в верхней трети бедра первые движения стопы и пальцев появляются лишь спустя 15–20 месяцев и позже после наложения нервного шва, а при ранениях этого же нерва в нижней трети бедра — через 10–15 месяцев после оперативного вмешательства. Наихудший прогноз отмечается при повреждениях на корешковом уровне, поскольку корешки спинальных нервов не регенерируют и не могут быть восстановлены хирургическим путем. Поражение корешков (обычно — отрыв корешка на шейном уровне), в отличие от поражения сплетения, характеризуется следующими признаками:
‒ интенсивная жгучая боль, иррадиирущая вдоль соответствующего дерматома;
‒ парализация паравертебральных мышц, иннервируемых задними ветвями спинальных нервов;
‒ паралич мышц лопатки вследствие нарушения функции коротких нервов плечевого пояса (крыловидная лопатка);
‒ синдром Горнера (при поражении С8-ты корешков);
‒ трофические нарушения и быстро прогрессирующая мышечная атрофия с грубыми вторичными контрактурами.
Периферические невропатии и плексопатии могут иметь самую различную этиологию. В мирное время наиболее частой формой поражений периферических нервов являются туннельные невропатии, составляя около 30–40 % от всех заболеваний периферической нервной системы. Туннельная невропатия — это локальное поражение нервного ствола, обусловленное его компрессией и ишемией в анатомических каналах (туннелях) или вследствие внешнего механического воздействия. Предрасполагающие к развитию туннельных невропатий факторы включают генетически обусловленную узость естественных вместилищ нерва, приобретенную узость этих вместилищ вследствие отеков и гиперплазии соединительной ткани при различных заболеваниях (например, сахарном диабете, гипотиреозе, коллагенозах), длительное перенапряжение мышечно-связочного аппарата у лиц определенных профессий, последствия трав, мышечно-тонические и нейро-дистрофические нарушения при рефлекторных синдромах остеохондроза позвоночника, ятрогенные травмирующие воздействия (неправильное наложение гипсовой повязки, кровоостанавливающего жгута). Нарушение функции нерва происходит вследствие как демиелинизации, так и поражения аксона (ухудшение нейротрофического контроля в связи с недостаточностью аксонального транспорта). Туннельные поражения нервов проявляются в первую очередь болью, чувствительными и вегетативными расстройствами. Двигательные нарушения развиваются лишь у одной трети больных и заключаются, как правило, в снижении мышечной силы, гипотрофии мышц, развитии контрактур. Прогноз восстановления функций при рано начатом лечении обычно благоприятный, однако это восстановление может происходить достаточно длительно, до нескольких месяцев. Кроме того, прогноз зависит от основного заболевания, на фоне которого развилась невропатия, от того, сохраняются ли профессиональные перегрузки конечности. В 30–40 % случаев туннельные невропатии рецидивируют.
На втором месте по частоте находятся травматические невропатии. Среди причин травматических невропатий прогностически наиболее благоприятными являются резаные ранения, при которых своевременное оперативное вмешательство обеспечивает хороший исход.
Тракционные и огнестрельные травмы имеют худший прогноз, поскольку при них нередко измененными оказываются центральный отрезок нерва и нейрон спинномозговых центров, что существенно затрудняет регенерацию нерва. Разрушение нервного ствола на большом протяжении может наблюдаться также при электротравме, при химическом повреждении (случайное введение в нерв различных лекарственных веществ). Весьма неблагоприятно сопутствующее повреждению нерва нарушение кровообращения в конечности (кровотечение либо длительное наложение кровоостанавливающего жгута, тромбоз магистральной артерии), которое может приводить к развитию в мышцах, сухожилиях, суставных сумках, коже и подкожной клетчатке атрофирующего склерозирующего процесса с образованием контрактур. Препятствовать восстановлению движений могут также вторичные изменения в суставах и сухожилиях, которые развиваются вследствие растяжения связок и суставных сумок при пассивном свешивании конечностей в случае вялого паралича или пареза. При невро- и плексопатиях, развившихся на фоне соматических заболеваний, вследствие иммунных, неопластических, инфекционных, токсических поражений и воздействий прогноз зависит от характера течения основного заболевания либо процесса.
Литература:
- Диагностика повреждения периферических нервов С. М. Рассел издательство: Бином Год: 2009 Страниц:251
- Неврология и нейрохирургия. Том 1. 4-е издание
- Автор: Гусев Е. И., Коновалов А. Н., Скворцова В. И.
- http://www.medicport.ru/doctors/stati_dlya_vrachej/materialy/lechenii_zabolevanij_perifericheskih_nervnyh_stvolov_i_spletenij/
- http://vse-zabolevaniya.ru/bolezni-nejrohirurgii/porazhenija-nervnoj-sistemy.html
- http://aupam.narod.ru/pages/medizina/reab_ruk_rbsdn_t2/page_28.htm
Основные термины (генерируются автоматически): нервный ствол, повреждение нерва, нерв, невропатия, гибель аксона, кровоостанавливающий жгут, миелиновая оболочка, нарушение функции нерва, основное заболевание, травматическая неврома.