Медицинская реабилитация
Основы нейронной теории
Нервная ткань образована двумя отличающимися морфологически и функционально типами клеток. К одному их них относятся собственно нервные клетки или нейроны, а к другому – клетки нейроглии или просто глии. И те, и другие происходят из общей популяции клеток-предшественниц, существующих только на ранней стадии эмбрионального развития мозга. В процессе дифференцировки два этих клеточных типа обособляются и затем специализируются на выполнении разных задач.
Путь к изучению морфологии т.е. внешнего вида, нейронов впервые нашёл в конце 60-х – начале 70-х годов XIX века итальянский врач и анатом Камилло Гольджи (Golgi С.). Однажды он решил использовать для окраски препаратов мозга нитрат серебра и обнаружил, что это вещество избирательно поглощается нервными клетками. В дальнейшем этот метод усовершенствовал знаменитый испанский гистолог Сантьяго Рамон-и-Кахал (Ramon y Cajal S.) и, пользуясь им, получил данные, которые позволили создать основы нейронной теории. В 1906 году Гольджи и Рамон-Кахал были награждены Нобелевской премией по физиологии и медицине «за работы по исследованию строения нервной системы».
Ещё в конце XVIII века итальянец Луиджи Гальвани (Galvani L.) обнаружил, что мышцы и нервные клетки животных производят электричество. В середине XIX века в Германии Эмиль Дюбуа-Реймон (DuBois-Reymond E.) уже располагал достаточно совершенной техникой для того, чтобы зарегистрировать биоэлектрические сигналы в нервном волокне. Герман Гельмгольц (von Helmholtz Н.) в 1854 году сумел измерить скорость прохождения нервных импульсов.
С той поры было выполнено огромное количество исследований структуры и функции нейронов, что это позволило сформулировать основы нейронной теории.
3.1. Нейроны
Среди 1011 нейронов человеческого мозга обнаружено множество непохожих друг на друга клеток, однако в их строении можно, как правило, найти общие отличительные признаки (Рис. 3.1). У каждого нейрона есть тело (другие названия этой части нейрона: сома, перикарион), где содержится ядро и цитоплазматические органеллы, где происходит синтез белков, нейромедиаторов и других важных компонентов жизнедеятельности клетки. При разрушении тела неизбежно гибнет и вся клетка.
От тела клетки отходят два типа отростков, представляющих собой тонкие нити цитоплазмы; они называются дендрит и аксон. Количество дендритов у различных клеток может существенно отличаться, у большинства нейронов их довольно много, причём каждый из дендритов ветвится подобно дереву, а его многочисленные ветви предназначены для приёма сигналов, передаваемых соседними клетками. Получив такие сигналы, дендриты проводят их к телу клетки.
В отличие от непостоянного количества дендритов у любой нервной клетки может быть только один аксон, который проводит электрические сигналы лишь в одном направлении: от тела клетки. Эти электрические сигналы называются потенциалами действия, они имеют амплитуду около 100 милливольт (мВ – тысячных долей вольта), а длительность около 1 миллисекунды (мс – тысячной доли секунды). Потенциалы действия обычно возникают в аксонном холмике – месте отхождения аксона от сомы и распространяются по аксону со скоростью от 1 до 100 м/с, не изменяя при этом амплитуду сигнала.
Диаметр аксона почти одинаков на всём протяжении, у разных клеток его величина варьирует от 0,2 до 20 мкм. Это обстоятельство сказывается на скорости проведения сигнала: чем толще аксон, тем быстрее проводится по нему потенциал действия. Длина аксонов у разных клеток может сильно отличаться: от 0,1 мм до 1 м (а у некоторых видов животных – до 3 м). Многие аксоны заключены в особый футляр, сформированный отростками некоторых клеток глии. Образует этот футляр миелин – жироподобное вещество со свойствами электрического изолятора: в области миелиновых покрытий электрические сигналы не возникают.
Миелиновая оболочка аксона регулярно прерывается участками, свободными от миелина – они называются перехватами Ранвье. По этим перехватам и распространяются потенциалы действия, как бы перескакивая через миелинизированные участки от одного перехвата к другому (такой тип передачи сигнала называется сальтаторным, от лат. saltare – прыгать), поэтому скорость проведения оказывается довольно высокой. Некоторые аксоны не имеют миелинового покрытия: в отличие от миелинизированных волокон их называют безмиелиновыми (по другой терминологии миелинизированные и безмиелиновые волокна различают как мякотные и безмякотные). По безмиелиновым волокнам потенциалы действия распространяется медленнее: здесь они не «прыгают», а «ползут» по всей длине аксона.
Природа потенциалов действия, с помощью которых передаётся информация в сенсорных, моторных или мотивационных системах, одна и та же, а сами потенциалы действия стереотипны. Их возникновение и проведение можно записать с помощью специальных приборов, но по характеру такой записи невозможно судить о том, какого рода информацию передают зарегистрированные потенциалы действия: о запахе ли цветущей сирени или о ласкающей глаз изумрудной зелени постриженного газона, о разгибании правой ноги в колене или о выделении слюны. Содержание передаваемой информации определяется не формой потенциалов действия, а специфическими объединениями нейронов в ту или иную сенсорную, моторную или мотивационную систему: каждый вид информации передаётся с помощью стереотипных потенциалов действия, но по собственному нейронному пути.
Неподалёку от своих окончаний большинство аксонов разделяется на тонкие коллатеральные ветви или аксонные терминали, причём некоторые из них могут даже повернуть назад – это обратные коллатерали. Терминали аксона вступают в контакт с другими клетками, чаще всего с их дендритами, реже – с телом и ещё реже – с аксоном. Аксоны эфферентных нейронов контактируют с клетками рабочих органов, которыми являются мышцы или железы внешней секреции. Контактная зона между двумя клетками получила название: синапс. В соответствии с этим термином клетка, передающая сигнал, называется пресинаптической, а получающая сигнал – постсинаптической. В подавляющем большинстве случаев эти клетки анатомически не соединяются и между ними находится синаптическая щель, которая заполнена жидкостью, напоминающей по своему составу плазму крови (особый вариант межклеточных контактов представляют электрические синапсы – см. главу 5).
Из-за анатомической разобщённости пресинаптическая клетка может повлиять на постсинаптическую только с помощью химического посредника – нейромедиатора или нейротрансмиттера. Медиатор должен выделиться из окончания аксона пресинаптической клетки тогда, когда к этому окончанию подойдёт потенциал действия.
3.2. Классификация нейронов
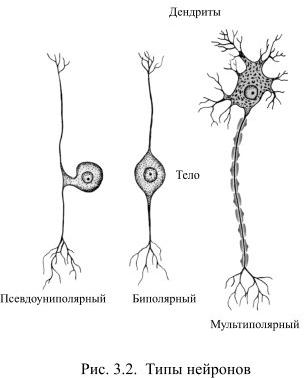
По количеству цитоплазматических отростков принято различать униполярные, биполярные и мультиполярные нейроны. Униполярные нейроны имеют единственный, обычно сильно разветвлённый первичный отросток. Одна из его ветвей функционирует как аксон, а остальные – как дендриты. Такие клетки часто встречаются в нервной системе беспозвоночных, а у позвоночных они обнаруживаются лишь в некоторых ганглиях вегетативной нервной системы.
У биполярных клеток есть два отростка (Рис. 3.2): дендрит проводит сигналы от периферии к телу клетки, а аксон передаёт информацию от тела клетки к другим нейронам. Так выглядят, например, некоторые сенсорные нейроны, встречающиеся в сетчатке глаза, в обонятельном эпителии.
К этой же разновидности нейронов следует отнести и чувствительные клетки спинальных ганглиев, воспринимающих, например, прикосновение к коже или боль, хотя формально от их тела отходит лишь один отросток, который разделяется на центральную и периферическую ветви. Такие клетки называют псевдоуниполярными, они формировались первоначально как биполярные нейроны, но в процессе развития два их отростка соединились в один, у которого одна ветвь функционирует как аксон, а другая – как дендрит.
У мультиполярных клеток один аксон, а дендритов может быть очень много, они отходят от тела клетки, а затем многократно делятся, образуя на своих ветвях многочисленные синапсы с другими нейронами. Так, например, на дендритах только одного мотонейрона спинного мозга образуется около 8000 синапсов, а на дендритах находящихся в коре мозжечка клеток Пуркинье может быть до 150 000 синапсов. Нейроны Пуркинье являются и самыми крупными клетками человеческого мозга: диаметр их тела около 80 мкм. А рядом с ними обнаруживаются крохотные зернистые клетки, их диаметр всего лишь 6-8 мкм. Мультиполярные нейроны встречаются в нервной системе чаще всего и среди них выявляется множество внешне не похожих друг на друга клеток.
Нейроны принято классифицировать не только по форме, но и по выполняемой функции, по их месту в цепи взаимодействующих клеток. Некоторые из них имеют специальные чувствительные окончания – рецепторы, которые возбуждаются при действии на них каких-либо физических или химических факторов, таких как, например, свет, давление, присоединение определённых молекул. После возбуждения рецепторов чувствительные нейроны передают информацию в центральную нервную систему, т.е. проводят сигналы центростремительно или афферентно (лат. afferens – приносящий).
Другая разновидность клеток передаёт команды от центральной нервной системы к скелетным или к гладким мышцам, к сердечной мышце или к железам внешней секреции. Это либо двигательные, либо вегетативные нейроны, по которым сигналы распространяются центробежно, а сами такие нейроны называются эфферентными (лат. efferens – выносящий).
Все остальные нейроны относятся к категории вставочных или интернейронов, которые образуют основную массу нервной системы – 99,98% от общего количества клеток. Среди них встречаются, как уже говорилось в главе 2, локальные и проекционные нейроны. Другое название проекционных нейронов – релейные; у них, как правило, длинные аксоны, с помощью которых эти клетки могут передавать переработанную информацию отдалённым регионам мозга. У локальных интернейронов аксоны короткие, эти клетки перерабатывают информацию в ограниченных локальных цепях и взаимодействуют преимущественно с соседними нейронами.
3.3. Электрические сигналы
Ещё Рамон-Кахал сформулировал два принципа, которые легли в основу нейронной теории и сохранили своё значение до настоящего времени:
1. Принцип динамической поляризации. Он означает, что электрический сигнал распространяется по нейрону только в одном и предсказуемом направлении.
2. Принцип специфичности соединений. В соответствии с этим принципом нейроны вступают в контакты не беспорядочно, но только с определёнными клетками-мишенями, причём цитоплазма контактирующих клеток не соединяется и между ними всегда сохраняется синаптическая щель.
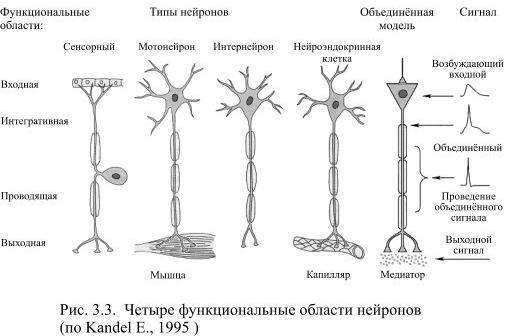
Современный вариант нейронной теории связывает определённые части нервной клетки с характером возникающих в них электрических сигналов. В типичном нейроне есть четыре определяемые морфологически области: дендриты, сома, аксон и пресинаптическое окончание аксона. При возбуждении нейрона в нём последовательно появляется четыре разновидности электрических сигналов: входной, объединённый, проводящийся и выходной (Рис. 3.3). Каждый из этих сигналов возникает только в определённой морфологической области.
Чтобы понять различия между этими сигналами, надо иметь некоторое представление о природе нервных импульсов. С наружной и внутренней стороны плазматической мембраны нейрона содержатся разные электрические заряды: с наружной стороны положительные, с внутренней – отрицательные. Разность между ними называется мембранным потенциалом покоя. Если считать наружный заряд равным нулю, то разность зарядов между наружной и внутренней поверхностями у большинства нейронов оказывается близкой к -65 мВ, хотя она и может у отдельных клеток варьировать от -40 до -80 мВ.
Возникновение этой разности зарядов обусловлено неодинаковым распределением ионов калия, натрия и хлора внутри клетки и снаружи её, а также большей проницаемостью покоящейся клеточной мембраны лишь для ионов калия (См. главу 4).
У возбудимых клеток, к числу которых принадлежат нервные и мышечные, потенциал покоя способен сильно изменяться и эта способность является основой для возникновения электрических сигналов. Уменьшение потенциала покоя, например, с -65 до -60 мВ, называется деполяризацией, а увеличение, например, с -65 до -70 мВ, – гиперполяризацией.
Если деполяризация достигнет некоторого критического уровня, например -55 мВ, то проницаемость мембраны для ионов натрия на короткое время становится максимальной, они устремляются в клетку и в связи с этим трансмембранная разность потенциалов стремительно уменьшается до 0, а затем приобретает положительное значение. Это обстоятельство приводит к закрытию натриевых каналов и стремительному выходу из клетки ионов калия через предназначенные только для них каналы: в результате восстанавливается первоначальная величина мембранного потенциала. Эти быстро происходящие изменения мембранного потенциала называются потенциалом действия. Потенциал действия является проводящимся электрическим сигналом, он быстро распространяется по мембране аксона до самого его окончания, причём нигде не меняет свою амплитуду.
Кроме потенциалов действия в нервной клетке, вследствие изменения её мембранной проницаемости, могут возникать местные или локальные сигналы: рецепторный потенциал и постсинаптический потенциал. Их амплитуда значительно меньше, чем у потенциала действия, кроме того, она существенно уменьшается при распространении сигнала. По этой причине местные потенциалы и не могут распространяться по мембране далеко от места своего возникновения.
3.4. Входные сигналы
Входными сигналами являются либо рецепторный, либо постсинаптический потенциал. Рецепторный потенциал образуется в окончаниях чувствительного нейрона, когда на них действует определённый стимул: растяжение, давление, свет, химическое вещество и т.п. Действие стимула вызывает открытие определённых ионных каналов мембраны, а последующий ток ионов через эти каналы изменяет первоначальное значение потенциала покоя; в большинстве случаев происходит деполяризация. Эта деполяризация и является рецепторным потенциалом, её амплитуда пропорциональна силе действующего стимула.
Рецепторный потенциал может распространяться от места действия стимула вдоль мембраны, но, как правило, на относительно небольшое расстояние. Дело в том, что амплитуда рецепторного потенциала уменьшается по мере удаления от места действия стимула и на расстоянии всего лишь 1 мм от этого места амплитуда рецепторного потенциала составляет только 1/3 от первоначальной величины, а ещё через 1 мм деполяризующий сдвиг и вовсе исчезнет.
Вторая разновидность входного сигнала – постсинаптический потенциал. Он образуется на постсинаптической клетке после того, как возбуждённая пресинаптическая клетка отправит для неё специального химического курьера – нейромедиатор. Добравшись путём диффузии до постсинаптической клетки, медиатор присоединяется к специфическим белкам-рецепторам её мембраны (См. главу 1), что вызывает открытие ионных каналов. Возникший в связи с этим ток ионов через постсинаптическую мембрану изменяет первоначальное значение потенциала покоя – этот сдвиг и является постсинаптическим потенциалом.
В одних синапсах такой сдвиг представляет собой деполяризацию и, если она достигнет критического уровня, то постсинаптический нейрон возбуждается. В других синапсах возникает противоположный по направленности сдвиг: постсинаптическая мембрана гиперполяризуется: величина мембранного потенциала становится больше и уменьшить её до критического уровня деполяризации становится труднее. Такую клетку трудно возбудить, она заторможена. Таким образом, деполяризующий постсинаптический потенциал является возбуждающим, а гиперполяризующий – тормозным. Соответственно этому и сами синапсы подразделяются на возбуждающие (вызывающие деполяризацию) и тормозные (вызывающие гиперполяризацию). Возбуждающие синапсы образуются, по большей части на дендритах постсинаптической клетки, а тормозные – на её теле.
Вне зависимости от того, что происходит на постсинаптической мембране: деполяризация или гиперполяризация, величина постсинаптических потенциалов всегда пропорциональна количеству подействовавших молекул медиатора, но обычно их амплитуда невелика. Так же, как и рецепторный потенциал, они распространяются вдоль мембраны на очень небольшое расстояние, т.е. тоже относятся к местным потенциалам.
Таким образом, входные сигналы представлены двумя разновидностями местных потенциалов, рецепторным и постсинаптическим, а возникают эти потенциалы в строго определённых областях нейрона: либо в чувствительных окончаниях, либо в синапсах. Чувствительные окончания принадлежат сенсорным нейронам, где рецепторный потенциал возникает под действием каких-либо раздражителей, внешних по отношению к нейрону стимулов. Для интернейронов, а также для эфферентных нейронов входным сигналом может быть только постсинаптический потенциал.
3.5. Объединённый сигнал – потенциал действия
Объединённый сигнал может возникнуть только в таком участке мембраны, где достаточно много ионных каналов для натрия. В этом отношении идеальным объектом является аксонный холмик – место отхождения аксона от тела клетки, поскольку именно здесь самая высокая во всей мембране плотность каналов для натрия. Такие каналы являются потенциалзависимыми, т.е. открываются лишь тогда, когда исходное значение потенциала покоя достигнет критического уровня. Типичное для среднестатистического нейрона значение потенциала покоя составляет приблизительно -65 мВ, а критический уровень деполяризации соответствует примерно -55 мВ. Стало быть, если удастся деполяризовать мембрану аксонного холмика с -65 мВ до -55 мВ, то там возникнет потенциал действия.
Деполяризовать мембрану способны входные сигналы, т.е. либо постсинаптические потенциалы, либо рецепторные. В случае рецепторных потенциалов местом возникновения объединённого сигнала является ближайший к чувствительным окончаниям перехват Ранвье, где наиболее вероятна деполяризация до критического уровня. В связи с этим надо учесть, что каждый чувствительный нейрон имеет множество окончаний, являющихся ветвями одного отростка. И, если в каждом из этих окончаний при действии стимула возникает очень небольшой по амплитуде рецепторный потенциал и распространяется к перехвату Ранвье с уменьшением амплитуды, то он является лишь малой частью общего деполяризующего сдвига. От каждого чувствительного окончания в одно и то же время перемещаются к ближайшему перехвату Ранвье эти небольшие рецепторные потенциалы, а в области перехвата все они суммируются. Если общая сумма деполяризующего сдвига будет достаточной, то в перехвате возникнет потенциал действия.
Постсинаптические потенциалы, возникающие на дендритах, так же невелики, как и рецепторные потенциалы и так же уменьшаются при распространении от синапса до аксонного холмика, где может возникнуть потенциал действия. Кроме того, на пути распространения постсинаптических потенциалов по телу клетки могут оказаться тормозные гиперполяризующие синапсы и потому возможность деполяризации мембраны аксонного холмика на 10 мВ кажется маловероятной. Тем не менее, этот результат регулярно достигается в результате суммации множества небольших постсинаптических потенциалов, возникающих одновременно в многочисленных синапсах, образованных дендритами нейрона с окончаниями аксонов пресинаптических клеток.
Таким образом, объединённый сигнал возникает, как правило, вследствие суммации одновременно образовавшихся многочисленных местных потенциалов. Такая суммация происходит в том месте, где особенно много потенциалзависимых каналов и поэтому легче достигается критический уровень деполяризации. В случае интеграции постсинаптических потенциалов таким местом является аксонный холмик, а суммация рецепторных потенциалов происходит в ближайшем от чувствительных окончаний перехвате Ранвье (или близко расположенным к ним участком немиелинизированного аксона). Область возникновения объединённого сигнала называется интегративной или триггерной (от англ. trigger – спусковой крючок).
Английский термин удачен своей метафорической выразительностью, так как накопление небольших деполяризующих сдвигов молниеносно трансформируется в интегративной зоне в потенциал действия, который является максимальным электрическим потенциалом клетки и возникает по принципу «всё или ничего». Это правило надо понимать так, что деполяризация ниже критического уровня не приносит никакого результата, а при достижении этого уровня всегда, независимо от силы стимулов, обнаруживается максимальный ответ: третьего не дано.
3.6. Проведение потенциала действия
Как уже говорилось, амплитуда входных сигналов пропорциональна силе подействовавшего стимула или количеству выделившегося в синапсе нейромедиатора – такие сигналы называют градуальными. Их длительность определяется длительностью стимула или присутствия медиатора в синаптической щели. Амплитуда и длительность потенциала действия от этих факторов не зависят: оба этих параметра всецело определяются свойствами самой клетки. Стало быть, любая комбинация входных сигналов, любой вариант суммации, при единственном условии деполяризации мембраны до критического значения, вызывает один и тот же стандартный образец потенциала действия в триггерной зоне. Он всегда имеет максимальную для данной клетки амплитуду и примерно одинаковую длительность, сколько бы раз ни повторялись вызывающие его условия.
Возникнув в интегративной зоне, потенциал действия быстро распространяется по мембране аксона. Это происходит благодаря появлению локального электрического тока. Поскольку деполяризованный участок мембраны оказывается иначе заряженным, чем соседствующий с ним, между полярно заряженными участками мембраны возникает электрический ток. Под действием этого локального тока деполяризуется до критического уровня соседний участок, что вызывает появление потенциала действия и в нём. В случае миелинизированного аксона таким соседним участком мембраны является ближайший к триггерной зоне перехват Ранвье, затем следующий, и потенциал действия начинает «перепрыгивать» от одного перехвата к другому со скоростью, достигающей 100 м/с.
Разные нейроны могут многим отличаться друг от друга, но возникающие в них потенциалы действия различить очень трудно, а в большинстве случаев и невозможно. Это в высшей степени стереотипный сигнал у самых разных клеток: сенсорных, интернейронов, моторных. Эта стереотипия свидетельствует о том, что сам потенциал действия не содержит никаких сведений о природе породившего его стимула. О силе стимула свидетельствует частота возникающих потенциалов действия, а определением природы стимула занимаются специфические рецепторы и хорошо упорядоченные межнейронные связи.
Таким образом, возникший в триггерной зоне потенциал действия быстро распространяется по ходу аксона к его окончанию. Это передвижение связано с образованием локальных электрических токов, под влиянием которых потенциал действия как бы заново возникает в соседнем участке аксона. Параметры потенциала действия при проведении по аксону нисколько не меняются, что позволяет передавать информацию без искажений. Если аксоны нескольких нейронов оказываются в общем пучке волокон, то по каждому из них возбуждение распространяется изолированно.
3.7. Выходной сигнал
Выходной сигнал адресуется другой клетке или одновременно нескольким клеткам и в подавляющем большинстве случаев представляет собой выделение химического посредника – нейротрансмиттера или медиатора. В пресинаптических окончаниях аксона заранее запасённый медиатор хранится в синаптических пузырьках, которые накапливаются в специальных участках – активных зонах. Когда потенциал действия добирается до пресинаптического окончания, содержимое синаптических пузырьков путём экзоцитоза опорожняется в синаптическую щель.
Химическими посредниками передачи информации могут служить разные вещества: небольшие молекулы, как, например, ацетилхолин или глутамат, либо достаточно крупные молекулы пептидов – все они специально синтезируются в нейроне для передачи сигнала. Попав в синаптическую щель, медиатор диффундирует к постсинаптической мембране и присоединяется к её рецепторам. В результате связи рецепторов с медиатором изменяется ионный ток через каналы постсинаптической мембраны, а это приводит к изменению значения потенциала покоя постсинаптической клетки, т.е. в ней возникает входной сигнал – в данном случае постсинаптический потенциал.
Таким образом, почти в каждом нейроне, независимо от его величины, формы и занимаемой в цепи нейронов позиции, можно обнаружить 4 функциональные области: локальную рецептивную зону, интегративную, зону проведения сигнала и выходную или секреторную зону (Рис. 3.3).
3.8. Глия
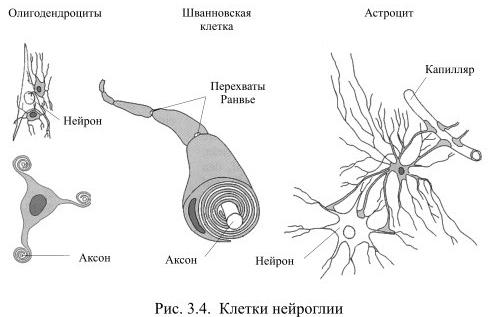
Во всех органах человеческого тела, кроме мозга, функционирующие клетки удерживаются вместе межклеточным веществом соединительной ткани. В нервной системе эту роль выполняет глия (от греч. глия – клей), клетки которой образуются из общих с нейронами предшественниц на раннем этапе развития мозга. Глия создаёт опору для нейронов, объединяет отдельные элементы нервной системы, но, в то же время, изолирует друг от друга разные группы нейронов, а также большую часть их аксонов. Тем самым она формирует структуру мозга. Численность клеток глии превышает количество нейронов в мозгу приблизительно в 10 раз. Эти клетки отличаются друг от друга по внешнему виду и по выполняемой функции (Рис. 3.4).
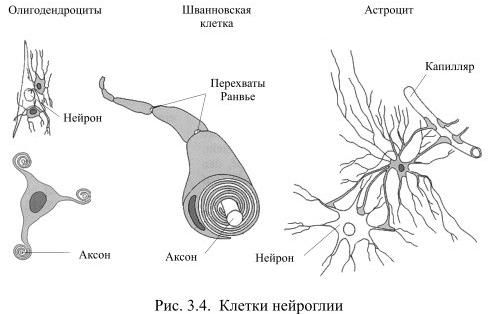
Самыми распространёнными среди клеток глии являются астроциты, например, в мозолистом теле они составляют 1/4 всех клеток глии. У астроцита неправильной, звёздчатой формы тело с многочисленными и относительно длинными отростками, одни из которых направлены к нейронам, а другие – к кровеносным капиллярам. Эти отростки расширяются на концах, образуя т.н. астроцитарную ножку. На поверхности капилляра отростки соседних астроцитов плотно смыкаются друг с другом и практически полностью обвёртывают кровеносный сосуд. Подобная изоляция сосуда является одним из способов формирования гематоэнцефалического барьера – границы между кровью и нервной тканью, закрытой для многих находящихся в крови веществ.
Другие отростки астроцита почти целиком обёртывают тела нейронов. Если нейрон возбуждается длительно, вокруг него повышается концентрация ионов калия, а это может уменьшить возбудимость соседних нейронов. Астроциты предупреждают такую возможность, поглощая излишки калия, – тем самым они выполняют функцию буфера. Некоторые клетки глии при этом деполяризуются, а поскольку они связаны между собою щелевыми контактами, между деполяризованными и находящимися в покое клетками возникает ток. Это, однако, не приводит к возбуждению, так как в мембране клеток глии очень мало потенциалзависимых каналов для натрия или кальция. Несмотря на то, что повышение концентрации ионов калия у астроцитов изменяет некоторые их свойства, в настоящее время нет достаточных оснований считать их прямыми участниками переноса нервных импульсов.
Две другие разновидности клеток глии: олигодендроциты и шванновские клетки похожи друг на друга по внешнему виду и по выполняемой функции. У них маленькое тело и относительно небольшие, как бы расплющенные отростки, которые многократно обёртывают аксоны нейронов, тем самым обеспечивая им изолирующий миелиновый футляр. Миелин – это жироподобное вещество, которое выполняет роль электроизолятора. При утрате миелиновой оболочки вследствие, например, демиелинизирующих заболеваний, передача сигналов из одной части мозга в другую серьёзно нарушается, что обычно приводит к инвалидности.
Олигодендроциты создают миелиновую изоляцию аксонов в центральной нервной системе, причём каждый олигодендроцит обслуживает, как правило, несколько аксонов. Шванновские клетки покрывают миелином волокна периферической нервной системы, причём каждая шванновская клетка занимается только одним аксоном.
В белом и сером веществе мозга рассеяны клетки микроглии. В отличие от других клеток глии в мозгу они – чужаки, пришельцы. Они образуются из моноцитов крови, сумевших пройти сквозь стенки капилляров в мозг, чтобы в нём поселиться (в других тканях такие оседлые моноциты называются макрофагами). Подобно макрофагам иных тканей клетки микроглии выполняют роль мусорщиков: они захватывают и разрушают обломки разрушающихся клеток, эта работа становится особенно заметной на фоне повреждений мозга.
Особую роль клетки глии выполняют, по-видимому, во время развития мозга. Некоторые их разновидности регулируют направление перемещения нейронов в определённые регионы растущего мозга, а также направление роста аксонов. Другие клетки глии возможно участвуют в питании нервных клеток путём регуляции кровотока, а тем самым транспорта глюкозы и кислорода.
Резюме
В выдающемся разнообразии индивидуальных признаков отдельных нейронов обнаруживаются общие черты, которые позволяют классифицировать нервные клетки по их строению и выполняемой функции. Электрические сигналы распространяются по нейрону только в одном направлении. В каждом нейроне можно выделить четыре морфологические области, выполняющие разные функциональные задачи. В каждой из этих областей возникает особая разновидность сигналов, используемых для передачи информации. Клетки глии, так же, как и нейроны, различаются по своему строению и выполняемой функции.
Вопросы для самоконтроля
31. Под буквами А-Д обозначена различная величина диаметра аксонов: по какому из них возбуждение должно распространяться быстрее?
А. 0,5 мкм; Б. 1 мкм; В. 3 мкм; Г. 6 мкм; Д. 9 мкм.
32. С какой частью постсинаптической клетки чаще всего вступают в контакт терминали аксона пресинаптической клетки?
А. Тело; Б. Сома; В. Перикарион; Г. Дендриты; Д. Аксон.
33. Какой из указанных ниже нейронов является эфферентным?
А. Передающий информацию о боли; Б. Передающий информацию о прикосновении к коже; В. Передающий информацию от скелетных мышц; Г. Передающий информацию от гладких мышц; Д. Передающий информацию железе.
34. Какая морфологическая область клетки чаще всего служит местом возникновения входного сигнала?
А. Дендриты; Б. Тело; В. Аксонный холмик; Г. Аксон; Д. Окончания аксона.
35. Что представляет собой рецепторный потенциал?
А. Входной сигнал; Б. Проводящийся сигнал; В. Объединённый сигнал; Г. Постсинаптический потенциал; Д. Выходной сигнал.
36. Что представляет собой потенциал действия?
А. Входной сигнал; Б. Объединённый сигнал; В. Выходной сигнал; Г. Местный потенциал; Д. Постсинаптический потенциал.
37. Какой сигнал возникает по правилу «всё или ничего»?
А. Входной; Б. Объединённый; В. Выходной; Г. Постсинаптический; Д. Местный.
38. Какой из указанных ниже сигналов является градуальным?
А. Потенциал действия; Б. Проводящийся; В. Постсинаптический; Г. Выходной; Д. Объединённый.
39. Какой сигнал возникает в триггерной зоне?
А. Постсинаптический; Б. Рецепторный; В. Входной; Г. Объединённый; Д. Выходной.
40. Какой из указанных ниже сигналов имеет наибольшую амплитуду?
А. Рецепторный; Б. Потенциал действия; В. Постсинаптический; Г. Местный; Д. Входной.
41. Чем непосредственно обеспечивается проведение сигнала по аксону?
А. Действием раздражителя; Б. Выделением нейротрансмиттера; В. Наличием миелинового покрытия; Г. Отсутствием миелинового покрытия; Д. Локальным электрическим током.
42. Что из перечисленного ниже имеет отношение к формированию гематоэнцефалического барьера?
А. Все клетки глии; Б. Астроциты; В. Олигодендроциты; Г. Шванновские клетки; Д. Микроглия.
43. Какие клетки осуществляют миелиновую изоляцию аксонов клеток центральной нервной системы?
А. Астроциты; Б. Шванновские клетки; В. Олигодендроциты; Г. Клетки микроглии; Д. Все клетки глии.
44. Какую функцию выполняет миелин?
А. Обёртывает тела нейронов, обеспечивая им механическую защиту; Б. Обёртывает кровеносные сосуды, создавая гематоэнцефалический барьер; В. Поглощает избыток ионов калия и тем самым выполняет роль буфера; Г. Является электроизолятором для аксонов; Д. Является проводником электрических сигналов.
45. При сдвиге значения мембранного потенциала до критического уровня должен возникнуть:
А. Потенциал действия; Б. Рецепторный потенциал; В. Постсинаптический потенциал; Г. Градуальный потенциал; Д. Входной сигнал.
Тест на тему «Физиология
клетки»
1.
Наибольшей возбудимостью обладают
клетки
-
эпителиальной
ткани; 2. неравной; 3. мышечной;
4.соединительной; 5. нет правильного
ответа -
Элементарные
единицы нуклеиновых кислот –-
аминокислоты;
2.нуклеотиды; 3. липиды; 4. полипептиды;
5. ионы кальция
-
-
В состав клеточной
мембраны входят
-
аминокислоты; 2.
нуклеиновые кислоты; 3. ионы железа; 4.
фосфолипиды; 5. рибосомы
-
Путем диффузии
через клеточную мембрану не могут
пройти
1. Углекислый газ; 2.Кислород; 3.
Полипептиды; 4. Витамины; 5. Жирорастворимые
гормоны
5. В
нервной ткани обнаружено разнообразных
молекул и-РНК
1. 100000; 2. около 200000; 3. 50000; 4. 20000; 5.
1000
6. В
состав ДНК не входит
1. Тимин; 2. Гуанин; 3. Цитозин; 4. Урацил;
5. Аденин
7.
Нитрат серебра при окраске комплекса
Гольджи способны удерживать
1.
протофибриллы; 2. микротрубочки; 3.
нейрофиламенты; 4. микрофиламенты;
5. глобулярные белки
8.
Энергетическими станциями клетки
являются
1. рибосомы; 2. митохондрии; 3. лизосомы;
4.митохондрии; 5. пластиды
9.
Форму клетки при сокращении способны
изменять
1.
Глобулярные белки; 2. Микрофиламенты;
3. Нейрофиламенты; 4. Протофибриллы; 5.
Нуклеиновые кислоты
10.
Быстрый транспорт веществ в клетке
осуществляется
1. Глобулярными белками; 2. Микротрубочками;
3. Нейрофиламентами; 4.Протофибриллами;
5.Микрофиламентами
Тест на тему «Физиология возбудимых тканей»
1. Терминали аксона
пресинаптической клетки чаще вступают
в контакт с
1.
телом; 2. сомой; 3. перикарионом; 4.
дендритами
2.Эфферентным является
нейрон
1.
передающий информацию о боли; 2.передающий
информацию о прикосновении к коже;
3.передающий информацию от скелетных
мышц; 4. передающий информацию железе
3. Местом возникновения
входного сигнала чаще служит
1.
дендриты; 2. тело; 3. аксонный холмик; 4.
аксон
4. Рецепторный
потенциал представляет собой
1.входной
сигнал; 2.проводящийся сигнал; 3.
объединённый сигнал; 4. постсинаптический
потенциал
5. Потенциал действия
– это
1.
входной сигнал; 2. объединённый сигнал;
3. выходной сигнал; 4. местный потенциал
6. По принципу «всё
или ничего» возникает сигнал
1.входной;
2. объединённый; 3. выходной; 4.
постсинаптический
7. Функция миелина
—
1.
обёртывает тела нейронов, обеспечивая
им механическую защиту
2.
обёртывает кровеносные сосуды, создавая
гематоэнцефалический барьер; 3. поглощает
избыток ионов калия и тем самым выполняет
роль буфера; 4. является электроизолятором
для аксонов
8. При сдвиге значения
мембранного потенциала до критического
уровня должен возникнуть
1.
потенциал действия; 2. рецепторный
потенциал; 3. постсинаптический потенциал;
4. градуальный потенциал
9. В клетке значительно
выше концентрация ионов
1.
натрия; 2. калия; 3. кальция; 4. хлора
10. При физиологическом
покое клетки открыты каналы для ионов
1.
всех катионов; 2. анионов; 3. натрия; 4.
калия
11. Величина равновесного
потенциала мембраны гигантского аксона
кальмара для ионов калия
1.
+55 мВ; 2. +25-30 мВ; 3. -60 мВ; 4. -75 мВ
12. Натрий-калиевый
насос считается электрогенным, поскольку
1. он
расходует энергию АТФ; 2. он создаёт
концентрационный градиент калия; 3. он
выносит из клетки натрий; 4. за один цикл
он удаляет из клетки положительный
заряд
13. Электрическое
поле между внутренней и наружной
поверхностями мембраны препятствует
вхождению в клетку
1.
калия; 2. натрия; 3. хлора; 4. кальция
14. Для состояния
рефрактерности характерно
1.
активированное состояние потенциалзависимых
каналов; 2. инактивированное состояние
потенциалзависимых каналов; 3. открытое
состояние потенциалзависимых каналов;
4. закрытое состояние потенциалзависимых
каналов
15. Блокатором ионных
каналов для калия является
1.
тетраэтиламмоний; 2. тетродотоксин; 3.
батрахотоксин; 4. кураре
Тест по теме «Рефлексы. Механизм мышечного сокращения»
1. Не является рефлексом
1.
Мигание в ответ на раздражение роговицы
инородным телом; 2. Кашель, вызванный
попаданием инородного тела в дыхательные
пути; 3. Образование антител в ответ на
поступление в организм чужеродного
белка; 4. Одышка, вызванная тяжёлой
физической работой.
2. К центральной
нервной системе не относятся
1.
Тела афферентных нейронов; 2. Тела
мотонейронов; 3. Вставочные тормозные
нейроны; 4. Вставочные возбуждающие
нейроны
3. Могут отсутствовать
в рефлекторной дуге
1.
Рецепторы; 2. Интернейроны; 3. Эфферентные
нейроны; 4. Эффекторы.
4. Не является
эффектором в рефлекторном ответе
1.
Скелетная мышца; 2. Гладкая мышца; 3.
Слюнная железа; 4. Фолликулы щитовидной
железы.
5. Составной частью
нервного центра являются
1.
Рецепторы; 2. Афферентные нейроны; 3.
Вставочные нейроны; 4. Эффекторы.
6. Свойство нервного
центра, обеспечивающее возникновение
рефлекторного ответа при ритмической
стимуляции одного афферентного входа
подпороговыми раздражителями
1.
Синаптическая задержка; 2. Трансформация
ритма; 3. Последовательная суммация; 4.
Посттетаническая потенциация.
7. Одновременное
действие подпороговых раздражителей
на всю поверхность рецептивного поля
называется
1.
Синаптическая задержка; 2. Трансформация
ритма; 3. Пространственная суммация; 4.
Последовательная суммация.
8. Вслед за ритмической
стимуляцией афферентного входа в нервный
центр рефлекса в течение некоторого
времени наблюдается повышенная
эффективность синаптической передачи.
Это может быть связано с
1.
Синаптической задержкой; 2. Трансформацией
ритма4 3. Пространственной суммацией;
4. Последовательной суммацией
9. Не является
обязательным для регуляции мышечного
напряжения
1.
Рецепторы Гольджи; 2. Афферентный нейрон;
3. Возбуждающий вставочный нейрон; 4.
Тормозной интернейрон
10. В ответ на лёгкий
удар неврологическим молоточком по
сухожилию четырёхглавой мышцы бедра
после короткого латентного периода она
сокращается и вследствие этого свободно
свисавшая голень приподнимается. Этот
рефлекс обусловлен раздражением
1.
Сухожильных рецепторов; 2. Рецепторов
Гольджи; 3. Осязательных рецепторов
кожи; 4. Проприорецепторов
11. Нечаянно дотронувшийся
до сильно нагретого предмета человек
немедленно отдёргивает от него руку.
Нервный центр этого рефлекса расположен
в
1.
Спинном мозге; 2. Стволе мозга; 3. Среднем
мозге; 4. Чувствительном ганглии
12. Цитоплазматическая
мембрана мышечного волокна называется
1.
Саркоплазма; 2. Сарколемма; 3.
Саркоплазматический ретикулум;
4.
Миофибрилла.
13. В миофибрилле
прикреплены к Z-мембране
нити
1.
Тропонина; 2. Тропомиозина; 3. Актина; 4.
Миозина
14. В образовании поперечных
мостиков при сокращении мышечного
волокна участвуют
1.
Тропонин – тропомиозин; 2. Тропонин-
актин; 3. Тропомиозин – актин;
4.
Миозин — актин
15. Участок миофибриллы
между соседними Z-мембранами
1.
Актиновый диск; 2. Миозиновый диск; 3.
Саркоплазма; 4. Саркомер.
16. Сократительный
белок
1.
Миозин; 2. Тропонин; 3. Саркомер; 4.
Сарколемма.
17. Сократительный
белок
1.
Актин; 2. Тропомиозин; 3. Миофиламент; 4.
Саркомер.
18. Рабочий орган
1.
Нервно-мышечный синапс; 2. Миофибрилла;
3. Двигательная единица;
4.
Эффектор.
19 К сокращению
мышечного волокна приводит
1.
Сокращение нитей актина; 2. Сокращение
нитей миозина; 3. Совместное сокращение
нитей миозина и актина; 4. Скольжение
нитей актина и миозина относительно
друг друга
20. В начале мышечного
сокращения происходит
1.
Присоединение ионов кальция к молекулам
тропонина; 2. Повышение концентрации
ионов кальция в межфибриллярном
пространстве; 3. Перемещение тропомиозина
в глубину желобка между нитями актина;
4. Прикрепление миозиновых головок к
актину
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
From Wikipedia, the free encyclopedia

Activity at an axon terminal: Neuron A is transmitting a signal at the axon terminal to neuron B (receiving). Features: 1. Mitochondrion. 2. Synaptic vesicle with neurotransmitters. 3. Autoreceptor. 4. Synapse with neurotransmitter released (serotonin). 5.Postsynaptic receptors activated by neurotransmitter (induction of a postsynaptic potential). 6. Calcium channel. 7. Exocytosis of a vesicle. 8. Recaptured neurotransmitter.
Axon terminals (also called synaptic boutons, terminal boutons, or end-feet) are distal terminations of the telodendria (branches) of an axon. An axon, also called a nerve fiber, is a long, slender projection of a nerve cell, or neuron, that conducts electrical impulses called action potentials away from the neuron’s cell body, or soma, in order to transmit those impulses to other neurons, muscle cells or glands.
Neurons are interconnected in complex arrangements, and use electrochemical signals and neurotransmitter chemicals to transmit impulses from one neuron to the next; axon terminals are separated from neighboring neurons by a small gap called a synapse, across which impulses are sent. The axon terminal, and the neuron from which it comes, is sometimes referred to as the «presynaptic» neuron.
Nerve impulse release[edit]
Neurotransmitters are packaged into synaptic vesicles that cluster beneath the axon terminal membrane on the presynaptic side of a synapse. The axonal terminals are specialized to release the neurotransmitters of the presynaptic cell.[1] The terminals release transmitter substances into a gap called the synaptic cleft between the terminals and the dendrites of the next neuron. The information is received by the dendrite receptors of the postsynaptic cell that are connected to it. Neurons don’t touch each other, but communicate across the synapse.[2]
The neurotransmitter molecule packages (vesicles) are created within the neuron, then travel down the axon to the distal axon terminal where they sit docked. Calcium ions then trigger a biochemical cascade which results in vesicles fusing with the presynaptic membrane and releasing their contents to the synaptic cleft within 180 µs of calcium entry.[3] Triggered by the binding of the calcium ions, the synaptic vesicle proteins begin to move apart, resulting in the creation of a fusion pore. The presence of the pore allows for the release of neurotransmitter into the synaptic cleft.[4][5] The process occurring at the axon terminal is exocytosis,[6] which a cell uses to exude secretory vesicles out of the cell membrane. These membrane-bound vesicles contain soluble proteins to be secreted to the extracellular environment, as well as membrane proteins and lipids that are sent to become components of the cell membrane. Exocytosis in neuronal chemical synapses is Ca2+ triggered and serves interneuronal signalling.[7]
Mapping activity[edit]
| Neuron |
|---|
|
Dendrite Soma Axon Nucleus Node of Axon terminal Schwann cell Myelin sheath |

Wade Regehr, a Professor of Neurobiology at Harvard Medical School’s Department of Neurobiology, developed a method to physiologically see the synaptic activity that occurs in the brain. A dye alters the fluorescence properties when attached to calcium. Using fluorescence-microscopy techniques calcium levels are detected, and therefore the influx of calcium in the presynaptic neuron.[8] Regehr’s laboratory specializes in pre-synaptic calcium dynamics which occurs at the axon terminals. Regehr studies the implication of calcium Ca2+ as it affects synaptic strength.[9][self-published source?][10] By studying the physiological process and mechanisms, a further understanding is made of neurological disorders such as epilepsy, schizophrenia and major depressive disorder, as well as memory and learning.[11][12]
See also[edit]
- Endoplasmic reticulum
- Golgi apparatus
- Micelle
- Membrane nanotube
- Endocytosis
- Vesicular monoamine transporter
Further reading[edit]
- Cragg SJ, Greenfield SA (August 1997). «Differential autoreceptor control of somatodendritic and axon terminal dopamine release in substantia nigra, ventral tegmental area, and striatum». The Journal of Neuroscience. 17 (15): 5738–46. doi:10.1523/JNEUROSCI.17-15-05738.1997. PMC 6573186. PMID 9221772.
- Vaquero CF, de la Villa P (October 1999). «Localisation of the GABA(C) receptors at the axon terminal of the rod bipolar cells of the mouse retina». Neuroscience Research. 35 (1): 1–7. doi:10.1016/S0168-0102(99)00050-4. PMID 10555158. S2CID 53189471.
- Roffler-Tarlov S, Beart PM, O’Gorman S, Sidman RL (May 1979). «Neurochemical and morphological consequences of axon terminal degeneration in cerebellar deep nuclei of mice with inherited Purkinje cell degeneration». Brain Research. 168 (1): 75–95. doi:10.1016/0006-8993(79)90129-X. PMID 455087. S2CID 19618884.
- Yagi T, Kaneko A (February 1988). «The axon terminal of goldfish retinal horizontal cells: a low membrane conductance measured in solitary preparations and its implication to the signal conduction from the soma». Journal of Neurophysiology. 59 (2): 482–94. doi:10.1152/jn.1988.59.2.482. PMID 3351572.
- LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite.[13]
References[edit]
- ^ «Axon Terminal». Medical Dictionary Online. Archived from the original on 2016-03-04. Retrieved February 6, 2013.
- ^ Foster, Sally. «Axon Terminal — Synaptic Vesicle — Neurotransmitter». Retrieved February 6, 2013.
- ^ Llinás R, Steinberg IZ, Walton K (March 1981). «Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse». Biophysical Journal. 33 (3): 323–51. Bibcode:1981BpJ….33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850.
- ^ Carlson, 2007, p.56
- ^ Chudler EH (November 24, 2011). «Neuroscience for kids Neurotransmitters and Neuroactive Peptides». Archived from the original on December 18, 2008. Retrieved February 6, 2013.
- ^ Rizo, Josep (2018-07-10). «Mechanism of neurotransmitter release coming into focus». Protein Science (Review). 27 (8): 1364–1391. doi:10.1002/pro.3445. ISSN 0961-8368. PMC 6153415. PMID 29893445.
Research for three decades and major recent advances have provided crucial insights into how neurotransmitters are released by Ca2+ -triggered synaptic vesicle exocytosis, leading to reconstitution of basic steps that underlie Ca2+ -dependent membrane fusion and yielding a model that assigns defined functions for central components of the release machinery.
- ^ Südhof TC, Rizo J (December 2011). «Synaptic vesicle exocytosis». Cold Spring Harbor Perspectives in Biology. 3 (12): a005637. doi:10.1101/cshperspect.a005637. PMC 3225952. PMID 22026965.
- ^
Sauber C. «Focus October 20-Neurobiology VISUALIZING THE SYNAPTIC CONNECTION». Archived from the original on 2006-09-01. Retrieved July 3, 2013. - ^
Regehr W (1999–2008). «Wade Regehr, Ph.D.» Archived from the original on February 18, 2010. Retrieved July 3, 2013. - ^ President and Fellows of Harvard College (2008). «The Neurobiology Department at Harvard Medical School». Archived from the original on 20 December 2008. Retrieved July 3, 2013.
- ^ «NINDS Announces New Javits Neuroscience Investigator Awardees» (Press release). National Institute of Neurological Disorders and Stroke. May 4, 2005. Archived from the original on January 17, 2009. Retrieved February 6, 2013.
- ^ «Scholar Awards». The McKnight Endowment Fund for Neuroscience. Archived from the original on 2004-05-08. Retrieved July 3, 2013.
- ^ Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D (November 1999). «LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite». Nature. 402 (6760): 421–5. Bibcode:1999Natur.402..421T. doi:10.1038/46574. PMID 10586883. S2CID 205056308.